Evolution in the Information Age: Rediscovering the Nature of the Organism
Department of Zoology
University of Toronto
Toronto, Ontario M5S 1A1
Canada
Abstract
The newest synthesis of evolutionary thought is emerging, and promises to return evolutionary biology to Darwin’s panoramic view of life. The key element is a long-standing dualism in evolutionary theory. This dualism has a long history within evolutionary biology, being manifested under guises such as: (1) the nature of the organism and the nature of the conditions, (2) internal and external, or intrinsic and extrinsic, factors, (3) production and exchanges, (4) boundary and initial conditions, (5) metabolism and replication, (6) energy and information, and (7) costs and benefits, and conflict and resolution. A partially retrospective review suggests that there is now a conceptual coherent framework for resolving the dualism, not by eliminating one component of the dualism but by integrating both.
Keywords: Evolution, Information, Evolutionary Transitions, Natural Selection, Environmental Selection, Sexual Selection.
![]()
For almost
150 years, the unifying principle of biology has been that evolution has
occurred. And yet, precisely how evolution occurs, and what, if any, are the
general principles of the major transitions in evolution, still remains a focus
of intense interest and scrutiny. Part of the problem stems from the basic
nature of living systems, which are simultaneously “in the environment” and
“part of the environment”. Darwin made this dual nature the cornerstone of his
views about evolution
...there are two factors: namely, the nature of the organism and the nature of the conditions. The former seems to be much more the important; for nearly similar variations sometimes arise under, as far as we can judge, dissimilar conditions; and, on the other hand, dissimilar variations arise under conditions which appear to be nearly uniform. Darwin (1872: 32)
Throughout its existence, evolutionary theory has oscillated between an over-emphasis on either the nature of the conditions or the nature of the organism, ranging from the debates between the “Darwinists” and the Weismannian “Neo-Darwinists” (Bowler, 1983) to those between the neo-Lamarckians and the saltationists and orthogenticists (Bowler, 1983) to the current debates between the “Panglossian adaptationists and constrained adaptationists” (Gould and Lewontin, 1979; Rose and Lauder, 1996) and the “functionalists and structuralists” (Goodwin, 1995).
I believe that
these debates have never provided definitive resolution because in each case
neither side has actually taken Darwin’s dictum seriously, and sought a true
synthesis of the nature of the organism and the nature of the conditions (though
presumably everyone involved felt that their particular viewpoint would
accomplish that goal). Evolutionary theory has thus lacked a significant
component of integration among processes derived from “the nature of the
organism” ("intrinsic" factors) and from the “nature of the
conditions” ("extrinsic" factors), operating on different temporal
and spatial scales (Brooks and Wiley 1988; Maynard Smith and Szathmary 1995,
1999). This is partly due to the lack of a common language or narrative encompassing
both aspects of the nature of life, and partly due to a lack of a common
conceptual framework and causal basis for such a common narrative. Finally, it is paradoxical that although its
foundational statement was titled Origin of Species, evolutionary theory has been under-developed with
respect to questions of the origins of transitions. A diffuse network of
thought that developed during the last quarter of the 20th century
provides such a framework. In this contribution I hope to outline the
fundamental elements of what I consider to be an emerging unified theory of
evolution.
1 The Nature Of The Organism Then And Now
1.1 Then: the Basic Units of Selection
Darwin thought that organisms were historically and developmentally cohesive wholes, and therefore it was in the "nature of the organism" to produce offspring that were all highly similar (but not identical) to each other and to their parents and other ancestors. He also postulated that reproduction produced variation without regard for environmental conditions and therefore it was in the "nature of the organism" to produce these offspring in numbers far exceeding the resources available for their support. When this inherent overproduction produced variety in critical characters, natural selection would preserve the versions that were functionally superior in that particular environmental context (adaptations). Whenever an environment changes, those organisms that already had the adaptations necessary to survive would do so, whereas those lacking appropriate adaptations would not. Selection did not create the adaptations, it only determined which ones, if any, would be favored for survival. The production of organismal diversity thus required that organisms be at once autonomous from, and sensitive to, the environment. Darwin's perspective contrasted sharply with Lamarck's proposal that adaptation was an immediate and directed response by organisms to their surroundings. Lamarck also believed that the nature of the organism was important in the production of diversity, but only because all organisms have the same ability to change according to their needs. So while Darwin postulated that the "nature of the organism" included autonomous, self-regulating properties, Lamarck believed that the "nature of the organism" was to be completely determined by the environment.
The distinction between Lamarckian adaptationism and Darwinian selectionism became increasingly blurred in the second half of the 20th century, as biologists focused more attention on parts of organisms and less on organisms as wholes. This reductionist movement, driven first by the successes of population genetics and later by the development of molecular methods, may have been an unconscious response to developmental biologists' discoveries of the complexities underlying the transition from DNA sequences to phenotypes; complexities that threatened to swamp simplistic theories based upon the mantra of one gene-one trait-one selection vector. Whatever the reasons, losing the perspective on whole organisms led to a loss of Darwin's panoramic view of biological diversity. In the last quarter of the 20th century there have been a number of efforts to re-emphasize the nature of the organism in evolutionary biology (e.g., Brooks and Wiley, 1988; Weber et al., 1988; Brooks et al., 1989; Brooks and McLennan, 1990, 1991, in press; Depew and Weber, 1995; Kampis, 1991, 1998; Kauffman, 1993; Maynard Smith and Szathmary, 1995, 1999; Odling-Schmee et al., 1996; Raff, 1996; Salthe, 1993; Van de Vijver et al., 1998; Brooks, 2000, 2001; Taborsky, 2000). These perspectives re-assert that orderliness and organization in biological systems result from the interaction of selection processes with the inherent nature of the organism.
1.2 The Nature Of The Organism Now: Metabolism And Information
The search for regularities in the behavior that characterizes the origin of transitions in natural systems can be approached from two perspectives, an "externalist" or "total system (TS)" perspective, in which emergent order in a subsystem of the total system is imposed by the rest of the system (the "surroundings"), and an "internalist" or "bound matter (BM)" perspective, in which emergent order results strictly from dynamical behavior of the system itself. Brooks and Wiley (1988) felt that evolution does not result from an extreme TS perspective nor from an extreme BM perspective, but from an interaction between a self-organized biological system (a BM component) and an organized environment (a TS component), each with their own "rules" of behavior. Nonetheless, most of the emphasis by Brooks and Wiley (1988) was on developing an understanding of the genealogical component of biological systems, leaving the impression that their proposal was an extreme BM stance. Others (notably Wicken, 1987 and Weber and Depew, 1995) have espoused a strongly TS perspective. This has led us back again into the old “internal/external” debate.
A fundamental basis of the proposal by Maynard Smith and Szathmary (1995, 1999) proposal is that organisms are both metabolic systems and information systems, and that many of the most important evolutionary transitions can be understood as trade-offs between the need to exist and the need to transmit information to the next generation. I believe this provides a suitable narrative framework for integrating two divergent viewpoints about the fundamental nature of organisms. Next I summarize efforts to formalize those perspectives.
1.3 Organisms As Metabolic Systems
Biological systems maintain themselves in highly organized states far from thermodynamic equilibrium with respect to their environments. Much has been written about this, little of which takes into account the constraining influence of accumulated genetic information on patterns of energy flow. Lotka (1913, 1925) was among the first 20th century authors to discuss biological systems in terms of energy flows and energy partitioning. He recognized that biological systems persist in space and time by transforming energy form one state to another in ways that that generate and maintain organized structure. Maurer and Brooks (1991; see also Brooks et al., 1989; Brooks and McLennan, 1990) recognized two classes of such energy transformations. Heat-generating transformations involve a net loss of energy available to the system, usually to energy in the form of heat. Conservative transformations involve changing free energy into states that can be stored and utilized in subsequent transformations; a fundamental example of this is the use of ATP to control the burning of glucose. Although all conservative transformations in biological systems are coupled with heat generating transformations, the reverse is not true; there is a heavy energetic cost to maintaining structure. Lotka (1913) suggested that the inevitable structural decay that must accompany such costs could be delayed, although not reversed, by the system's accumulation of bound energy from conservative transformations. Or, the interplay between flow and partitioning of energy in biological systems acts to slow the rate at which energy stored by conservative transformations is degraded by heat-generating transformations.
Entropy changes (dS) can be subdivided into two components, one measuring exchanges between the system and its surroundings (deS, observed as changes in the surroundings) and the other measuring production by irreversible processes internal to the system (diS, observed as changes within the system). Exchanges between biological systems and their surroundings are accompanied by a great deal of waste; hence, deS is very large compared with diS. However, if biological systems are able to maintain their structural integrity, they must produce entropy internally (diS > 0). Or,
dS = deS + diS, diS >0
Therefore, diS (internal production) is critically important in biological evolution, even though it represents a very small portion of the total energy budget for biological systems (Maurer and Brooks, 1991).
Production rules in biological systems are those processes for which there is an energetic "cost" or "allocation". Following Prigogine and Wiame (1946) and Zotin and co-workers (e.g., Zotina and Zotin, 1978; see also Gladyshev, 1996), Brooks and Wiley (1988) denoted such allocations using the symbol y, denoting a specific dissipation function. The function includes at least two major classes of processes: (1) those involved in dissipation from the system, called the external dissipation function (ya, e.g. thermal entropy) and (2) those involved in dissipation within the system, called the bound dissipation function (ym, or structural entropy). In biological systems, ym can be further subdivided into allocations for accumulating biomass (ymb) and allocations for accumulating genetic diversity (ymi). Brooks and Wiley (1988) suggested that all three components of biological production (y) should be included in the thermodynamic production term diS, shown heuristically as
diS
= ya
+ ymb
+ ym i
Energy used in the uptake of raw materials from the surroundings into the system produces entropy which is dissipated into the surroundings (entropy production resulting from exchanges between the system and its surroundings, deS: Prigogine and Wiame, 1946; Prigogine, 1980). Different manifestations of entropy production (entropy production resulting from irreversible processes within the system, diS: Prigogine and Wiame, 1946; Prigogine, 1980) can be associated with each of the classes of transformations. Heat-generating processes occur when energy and entropy flow in opposite directions, entropy production tending to move the system towards disordered states. Conservative transformations are characterized by energy and entropy flowing in the same direction, entropy production being retained within the system and tending to move the system towards more structured states. As entropy and energy flow through biological systems at different rates, structure accumulates at different levels of organization; furthermore, the structure at any given level is constrained by energy and entropy flows at other levels. Rate gradients in entropy production lead to different types of constraint systems, including phylogenetic constraints, governing hierarchically related entities.
Organisms and the genealogical systems they form are maintained through time by the exploitation of "entropy gradients" or "resource gradients" in the surroundings (Wicken, 1987; Ulanowicz, 1988, 1997; Matsuno, 1989, 1995, 1996, 1998, 2000; Hirata, 1993; Depew and Weber, 1995), determined by interactions between abiotic and biotic factors. Abiotic factors can be structured in part by the ya component of the genealogical hierarchy. For example, metabolic processes are involved in the degradation of high grade energy sources into lower grade forms of energy, including heat. Both the capture of incoming solar energy by biological systems, and the mass re-radiation of heat by these organisms affects the thermal profile of the earth. Additionally, the production of oxygen as a byproduct of photosynthesis or of carbon dioxide as a byproduct of aerobic metabolism affects the composition of the earth's atmosphere. This means that the production term (diS) can influence the exchange term (deS). Biotic factors are subject to the influences of the structural portion of the genealogical hierarchy (ymb + ym i). The ecological hierarchy has a propensity to move the products of the genealogical hierarchy in the direction of minimizing energy gradients in the environment, to the extent permitted by the inherited capabilities (and limitations) of the members of each species (Gladyshev, 1996; Ulanowicz, 1997; Brooks and McLennan, 2000).
Entropy is produced at different rates in biological systems because energy stored by conservative transformations is degraded at different rates. Thus, biological systems manifest organized structures on different temporal and spatial scales. At the lowest organizational levels, the shortest time intervals, and the smallest spatial scales, the greatest relative contribution to y will be ya. If we examine cellular or sub-cellular structure over short time intervals, processes such as metabolism and respiration dominate explanations of observed structure. Most entropy production is dissipated into metabolic heat loss, and biological systems behave as classical dissipative structures. At more intermediate levels of organization, space or time, the effects of ymb predominate. Most entropy production at this scale is dissipated into accumulation and maintenance of biomass. Finally, on the largest and longest scales, ym i predominates, and the patterns relevant to biological explanations are formed mainly by accumulation and maintenance of genetic diversity. From the perspective of the environment, such patterns of biodiversity tend to be organized with respect to energy gradients, whereas from the perspective of the genealogical system, biodiversity is organized with respect to sister-group relationships and patterns of geographical distribution that mirror geological evolution occurring on similar temporal and spatial scales.
From an energetic perspective, organisms have a
dualistic nature. As open thermodynamic systems, they must simultaneously
interact with their surroundings and perform critical autonomous functions
internally. They maintain themselves in a viable state by exchanging matter and
energy irreversibly with their surroundings, taking in relatively high grade
energy and using it to perform useful work within themselves. This requires
sensing of, and causal engagement with, the surroundings, mediated by a physical
distinction ("phase separation": Prigogine, 1980) between the
organism and its surroundings. That is, there must be an “inside” and an
“outside” of the organism, delineated by a physical boundary. For all
organisms, this boundary is provided by cell membranes, which are
simultaneously physical barriers between the inside and outside of the organism
and highly selective mechanisms for modulating the exchange of matter and
energy between the organism and its surroundings. For multicellular organisms, this
barrier is a complex of cell membranes.
As well, there must be an energetic “trade-off” between exchanges and production. The proportion of any organism's total energy budget allocated for transmitting information from one generation to the next is very small. Nonetheless, this tiny component is essential to understanding evolution and the generalities of historical uniqueness, cohesion, and functional integration. This component is so small compared to the total thermal entropy production of the system, most of which is lost from the system, that the entropy of the surroundings increases much more than the entropy of the system. Hence, biological systems remain low-entropy (far from equilibrium) systems relative to their surroundings. However, there is a net increase in entropy within the system, indicating that evolution in its most general (and thus least predictable for specific cases) sense results from the propensity for systems to become more entropic over time. In general, we would expect any changes increasing the efficiency of internal production to be favored evolutionarily. Maynard Smith and Szathmary (1995, 1999) stated that the major transitions in evolution all involved increased efficiency of storage and transmission of biological information.
1.4 Organisms As Information Systems
Elucidation of the genetic code made it possible to begin thinking of organisms as information systems. A keystone of the theoretical constructs proposed by Brooks and Wiley (1988) and by Maynard Smith and Szathmary (1995, 1999) is that organisms are information systems. This seemingly simple assertion has created substantial controversy, however.
1.4.1 The Controversy
Information theory has been developed from two general perspectives, "communications theory" and "physical measurement theory". In classical communications theory, the amount of information sent from a source is calculated using a statistical entropy function. Errors in transmission can result from poor encoding at the source or from noise in the transmission channel. The meaningful information is that subset of information transmitted which is actually recorded by the receiver (there may or may not be a separate decoder). All of the processes affecting the transmission and reception of the information thus decrease the entropy of the message from its maximal value at the source. Physical entropies are expected to increase as a result of work done on the system, so either information transmission is not a physical process or the communications view of entropy is non-physical. Physical measurement theory provides a second formalism for information. Brillouin (1962) distinguished "free information", an abstraction involved in descriptive exercises, and "bound information", referring to material properties of the system (but not stating that information is a material part of the system per se). Bound information is determined with respect to the "complexions" (microstates) of the system. Hence, it is also calculated using a statistical entropy function but, contrary to communications theory, is expected to exist only in systems for which there is a non-arbitrary microstate/macrostate distinction [i.e. I(M)= f(Mm)]. Bound information is defined as
I
= Hmax - Hobs
where Hmax refers to the totally relaxed state of the system (usually estimated by a randomization of the observed components of the system). Brillouin defined I as "negentropy", which is converted into bound information by measurement (measuring devices are receivers), so negentropy = information. Information thus has a physical basis, but is not a material part of the system.
There must be an additional conception of information for biology because biological information (based on nucleic acids) has both communications functions and a physico-chemical basis. The conception must also an account of the growth of information through time. Biologists do not think that all possible genotypes were present at the beginning of life, and that since then those possibilities have been reduced to the diversity that we see today. Our empirical evidence suggests that biological information has grown over time. What is needed is an account of biological information that is (1) physically realistic (can be shown to have an objective basis), (2) intrinsic to the system (is a material part of the system) rather than to devices for measuring the system, and (3) can grow spontaneously over time. The two basic issues with respect to information and entropy are (1) whether information can be a material part of a system rather than just an abstract representation and (2) whether or not there is an objective difference between macrostates and microstates in calculations of informational entropies.
1.4.2 A Proposed Solution
John Collier (1986, 1988, 1990, 1992, 1996, 1998, 2000) has proposed that biological information is related to concepts of the causal capacity of a system, or its ability to impose distinctions on its surroundings. In a way, the emphasis is on how the system produces effects on measuring devices and not on how the measuring devices are affected. Collier proposed that physical (=material) information systems occur as arrays, or multi-dimensional messages, in which macrostate and microstate distinctions are distinguished non-arbitrarily, and that in order for this information to be related to physical concepts there must be (1) a physical (material) basis for the information, (2) an energetic cost in producing the information, and (3) a real (non-arbitrary) macrostate/microstate distinction. Since the discovery of the chemical structure and function of DNA, there has been a material basis for biological information, satisfying (1) above (see also Collier and Hooker, 1999; Brooks et al., 1989; Smith, 1988, 1998, 2000).
Energy dissipated within the system as a result of work done on the system (including heat-generating transformations, or ya of diS) is intropy, which stands for internal entropy (overhead of Ulanowicz, 1986, 1988, 1997). Energy that is converted into structure (conservative transformations, or ym of diS) is enformation, standing for intrinsic information (these distinctions originated in Collier, 1990). Conservative processes within biological systems are coupled with heat-generating processes, so there is an energetic cost associated with the production and maintenance of biological information. Intropy and enformation are interconvertable (e.g., energy brought in from the surroundings can be converted into structure, say glycogen, which can then be converted into heat). Intropy is converted into enformation by cohesive properties of the system. Cohesive properties, ranging from molecular affinities to cell-cell adhesion to genetic compatibility, mate recognition, and genealogy, also provide resistance to fluctuations from lower levels, allowing macroscopic properties to emerge. Cohesion is thus analogous to inertia. The major transitions in evolution discussed by Maynard Smith and Szathmary (1995, 1999) are all associated with the emergence of new forms of cohesion, which permit information to be stored and transmitted more efficiently. It is in the recognition of the significance of cohesive properties that we find the key to understanding microstate/macrostate distinctions in biological systems.
Finally, macrostate/microstate distinctions are determined objectively by part/whole associations. The number of accessible microstates is increased by the production of new components, either at a given level or through the opening up of new levels of organization. Biological systems accomplish this by conservative transformations. For example, auto-catalytic processes producing monomers make "monomer space" available for molecular evolution. Some monomers have high chemical affinities for each other, and will spontaneously clump into dimers and polymers. Once polymers begin to form, "polymer space" becomes available to the evolving system. At this level, polymers are macrostates and monomer and dimer distributions are microstates. Causal interactions among polymers create new levels of organization in which polymer distributions are the microstates and new levels of organization are the macrostates, and so on. Each new functional level creates a hierarchy of increasing structural intricacy, manifested by increasing allocation of the entropy production in structure. Therefore, the allocation of diS to ym might be nearly proportional to entropy increases due to the expansion of phase space resulting from the creation of new possible microstates. A protein coding unit might be considered a macrostate, while all the actual sequences that code for that protein would be the microstates; a locus could be a macrostate, and all alleles corresponding to that locus the microstates; phenotypes could be macrostates, and all genotypes corresponding to a given phenotype would be microstates (Layzer, 1978, 1980; Collier, 1986; Brooks and Wiley, 1986, 1988; Kauffman, 1993). This formulation answers objections that biological informational entropies do not include a macrostate/microstate distinction; Wicken (1987) first noted that this was a critical issue to be resolved and it remains problematical for those who fail to appreciate the relevance of the “nature of the organism” (the genealogical information system and its hierarchical organization).
In
discussing the origin of life and the extent of biological diversity, Maynard
Smith and Szathmary (1995, 1999) noted that living systems are digital rather
than holistic replicators, and
suggested that this was a key element in understanding that living systems
exhibited essentially unlimited capacity for variation. The formalism I have
summarized above supports this assertion. Consider a replicator
comprising a string of DNA 1,000 bases long. Given 4 nucleotides, the string has at
most 2,000 bits (this fully defines its structure at all levels). If the string
were a holistic replicator, it would function strictly as a single unit. If
that string functioned as a digital replicator, however, it could be read at
multiple levels (in thise, 1,000 levels from single bases to the entire
sequence) to produce a diversity of information. Each of these readings has a
maximum information capacity of 2,000 bits. Thus, if each reading of a given
sequence is equivalent to all others informationally, and if these readings are
not interactive (i.e., not cohesive), the total possible readings have a
maximum information capacity of 2,000,000 bits (see also Brooks et al.,
1984, 1988; Smith, 1988, 1998, 2000). If the bases
are interactive, then these self-interactions will constrain the total
information capacity. The total amount of information that could be expressed
at any one point in time is highly constrained by the fact that bases (a
similar argument holds for genes, tissues, and organisms) are causally linked,
so accessing some information will limit (or eliminate) expression of other,
cohesion therefore putting an upper bound on the amount of information
potential/capacity that could be expressed at any one time. At the same time,
accessing the same system in different ways sequentially through time permits
the same constrained quantity of information to be additive, since at each
point in time it is a different 2,000 bits being expressed.
Phenotypic diversity at all levels, from organisms to ecosystems, is coupled with genetic conservatism. The realization of so many possibilities indicates that multiple readings occur (i.e., DNA functions digitally in living systems). As mentioned above, however, not all of the possibilities are realized, due to cohesive interactions among the various “building blocks”. How do the expressed multiple readings maintain sufficient independence so as not to collapse to the same 2000 bits of information repeated over and over again? One mechanism is temporal separation; different readings of the same sequences may occur sequentially in time during, e.g. a cell cycle, the ontogeny of a multicellular organism, or the breeding cycle of a population. Structural and functional cohesion may constrain the maximum quantity of expressed information per unit time, but over a time sequence, it is possible to express and accumulate various manifestations of the same overall quantity of information. We know that the same genome produces very different products at different times during ontogeny. We also know that the same genome can produce diverse outcomes in different parts of the organism during ontogeny; spatial separation thus functions as another mechanism allowing relative independence of readings. It does so by creating boundary effects, or the “context” within which the same or different readings function. Any given reading can at best contain the only distinctions in what is transcribed by the reading. It does not follow from this that what is read functions the same way in each “time” or each “place”, though, since the reading brings historical information (about both text and context) with it. The effects of priori readings may change the system enough to create a new context in which the same reading manifests different outcomes, or different readings may produce the same outcome. There is no obvious constraint on the possible functional distinctions, except that they must be passed on, which tends to filter out any circumstantial or accidental functionality through time.
Biological information thus grows through non-linear interactions that produce branching through both time and space. If the phenotype were merely a linear expression of the genotype, then there would be no new information. But the branching produces information (realized functional phenotypic diversity) not completely specified by the genes or by the environment. Or, to return to Darwin, (1) evolution is the result of interactions between the nature of the organism and the nature of the conditions, and (2) the nature of the organism is the most important component, because it is the nature of organisms to be digital replicators and it is digital capacity that permits evolution to be essentially unlimited.
1.5 Biological Information Through Time
Brooks and Wiley (1986, 1988) examined different kinds of biological processes that show direction in time, growth and increasing complexity, coupled variation and constraint, and hierarchical and self-organization, to see if there were any underlying generalities. They used a general mathematical formalism summarizing changes in the number of things, the number of kinds of things, and the relative frequency of the different kinds of things, which are the key elements of (at least) biological complexity. This simple informational "entropy function" has four potentially illuminating formulations: (1) the "actual" entropy (Hobs) calculated on the basis of the observed distribution of components of the system (also called the information content); (2) the "maximum possible" entropy (Hmax), estimated by calculating the entropy value for the components of the system at any given time if they were all randomized (also called the information capacity); (3) an absolute difference (Hmax-Hobs) (also called Information or Macroscopic Information: Gatlin, 1972, Layzer, 1975; Landsberg, 1984a,b); and (4) two conceptually related relative differences (Hobs/Hmax, and Id/Hmax, Id being the information density, called Order (Landsberg, 1984a,b) and Redundancy (Gatlin, 1972), respectively. Even simple heuristic simulations emulating biological processes associated with the storage and transmission of information (e.g., reproduction, ontogeny, and speciation) produce three generalities (Brooks and Wiley, 1986, 1988: summarized in Fig. 1): (1) Hobs is an increasing function of time; (2) Hobs is a concave function of time, as historical constraints retard the rate of entropy increases; and (3) the difference between Hmax and Hobs is an increasing function of time, permitting the growth of structure and organization (for details see Collier and Hooker, 1999). Thus, both entropy and information/organization/order (redundancy) increased over time (see also Brooks et al., 1984, 1988; Smith, 1988, 1998, 2000). If we view Hmax as a function of the capacity, or potential, of a system and Hobs as a function of the content, or expression, of some of that potential, the difference between total information capacity and information content could be interpreted as being a function of the constraints, both inherent (BM) and extrinsic (TS), placed on the system (Brooks and Wiley, 1988; Smith, 1988, 1998, 2000; Brooks et al., 1984, 1988, 1989; Brooks and McLennan, 1990, 1991, 2000; Brooks, 1991, 1992, 1994, 1997, 1998, 2000, 2001).
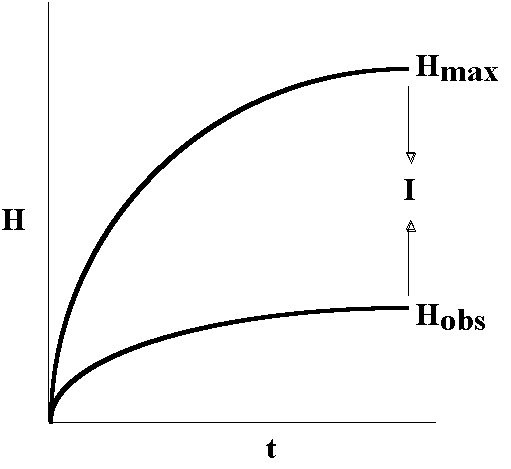
Figure 1. An heuristic view of information production in material systems.
Not all phenomena that can be characterized by this informational entropy function are bound by any regularities related to the Second Law of Thermodynamics. Does this behavior indicates anything causal, or if it is simply descriptive? That is, does any physical model explain these regularities to such an extent that we might say, in retrospect, that such regularities are expected? Frautschi (1982, 1988; also Layzer, 1975; Landsberg, 1984a,b) contrasted 2 classes of processes that generate entropy. The first is equilibration of temperatures between system and surroundings. Biological systems exhibit this kind of entropic behavior through processes that result in the dissipation of energy (heat loss) to their surroundings. The second is expansion of the phase space (i.e., the realm of all possibilities) in which the system resides, increasing its number of accessible microstates (possible configurations). System organization increases so long as equilibration (equiprobable distribution of the system over its microstates) occurs at a slower rate than the expansion of the phase space, allowing a lag between the increase in realized entropy (Hobs) and the increase in the maximum possible entropy of the system (Hmax), which is a linear function of the logarithm of the number of states or size of the phase space. So long as the phase space expands faster than the system can fill it up, increasing entropy can be accompanied by the emergence of organized structure. In cosmology, this argument explains the spontaneous and irreversible emergence of stars, solar systems, galaxies, and other organized structures, in which fundamental forces linking material bodies slow down the entropic diffusion of matter in the universe to such an extent that organized structures emerge as a result of and not at the expense of increasing entropy. A similar phenomenon accompanies the expansion of energy from the sun into space (Frautschi, 1982, 1988). In biological systems, mutations (as well as higher order genotypic and epigenetic phenomena) act to expand the genetic phase space within which life occurs, while genetic and genealogical bonds linking all living systems (their inheritance systems: Maynard smith and Szathmary, 1995, 1999), as well as the environments in which they exist, play roles analogous to the fundamental forces linking material bodies (Layzer, 1978, 1980; Brooks and Wiley, 1988; Smith, 1988, 1998, 2000; Brooks et al., 1984, 1988, 1989; Brooks and McLennan, 1990, 1991, 2000; Brooks, 1991, 1992, 1994, 1997, 1998; Kauffman, 1993; Kjellstrom, 1996; Lemke, 1998; Niklas, 1999; Shpak and Wagner, 2000).
Both biological and cosmological evolution exhibit long time scale regularities. Both show positive correlation between the spatial scale and the temporal scale of the emergence of regularities (e.g., Brooks, 1988; Maurer and Brooks, 1991). Both are also characterized by entropy production, which requires a nonequilibrium state, and by the emergence of low-entropy structures, which are the sources of continuing entropy production. This apparent paradoxical behavior occurs not as a result of the degradation of a static environment by the entropy production of the system [although biological systems do degrade their environments as a result of using energy and matter form their surroundings], leaving the system in a truly negentropic state, but from the behavior of a dynamic environment in which maximum possible entropy increases faster than the systems which produce the entropy can relax. In cosmological models, the dynamic environment is produced by the expansion of the universe and symmetry-breaking into distinct forms, such as matter and radiation. We characterize the resulting order as the energy in the universe available to do work.
A Unified View of the Nature of the Organism
Biological systems, beginning with organisms, are functional wholes with respect to the way they engage their surroundings as well as with respect to their internal organization. A major component of internal organization is functional integration through the interdependence of parts, and this is most evident in the dynamics of ontogeny (Salthe, 1993; Raff, 1996) and in physiological processes. Evolutionary changes in biological systems do not occur all at the same time; thus, when such changes occur, only part of the system changes. All changes, whether they are point mutations on a chromosome, genomic doubling, or alteration of part of a complex mating ritual, must integrate with the rest of the system, which has not changed, in such a way that viable organisms result. The functional necessity of developmental integration creates stability domains within bio-informational phase space (Brooks and Wiley, 1988; Kauffman, 1993; Niklas, 1999). Orderliness and organization in biological systems results from the interaction of selection processes with three aspects of the nature of the organism: (1) historical uniqueness, (2) cohesive properties, and (3) hierarchical organization.
The historical uniqueness of biological systems manifests itself in 2 ways. One is historical contingency; events that take place in the past may have an effect on the subsequent behavior and fate of the system. The other is temporal irreversibility; many biological processes are tied to the directional nature of time, albeit on different time scales. Some important biological processes, such as reproduction, development, aging and death, speciation, and extinction, appear to be temporally irreversible on any time scale. Until recently, the theory of natural selection has been generally considered a theory of reversible processes (e.g., Maynard Smith, 1970; but see Demetrius, 1992; Maynard Smith and Szathmary, 1995, 1999; Smith, 1998, 2000). Some evolutionary changes appear to occur on longer time scales than those provided by environmental selection. If some long-term irreversible behavior in evolution is not just a result of historical contingency, evolutionary theory needs a causal explanatory principle to account for it. Spontaneous irreversible behavior in biological systems always involves two things: (1) growth and increasing complexity (Brooks et al., 1989), and (2) physical manifestations of at least some of the systems' history (Wimsatt, 1988). Maynard Smith and Szathmary (1995, 1999) have suggested that the conjunction of two or more events, each on inherently improbable by itself, is sufficient to render a change irreversible. This perspective lends itself well to the above formulation of the dynamics of production of biological information, without sacrificing significant roles for historicity or selection.
Cohesive properties of biological systems range from cell-cell adhesion and recognition, to sexual reproduction and specific mate recognition systems, to common phylogenetic history. Cohesion is especially important to evolutionary explanations, because the cohesive properties of living systems limit the ways in which and the extent to which populations can respond to environmental selection, and are the "glue" of functional integration and hierarchical organization that are so characteristic of biological systems (Wake and Roth, 1989). Many biological processes that demonstrate irreversible behavior manifest such changes as a result of interactions among cohesive factors, which tend to keep species and populations together, and diversifying factors, which tend to split them into separate systems. Speciation in sexually reproducing species, for example, results when the developmental and reproductive constraints acting as cohesive forces maintaining an ancestral species as a single lineage are overridden by environmental forces acting to split it apart into descendant species (Wiley, 1981; Wiley and Brooks, 1982; Brooks and Wiley, 1986, 1988; Brooks and McLennan, 1991, in press); this is the reason Maynard Smith and Szathmary (1995, 1999) associated the origin of sex with the origin of true biological species. In a complementary fashion, the environmental boundaries within which each species lives might, in some cases, be pronounced enough to be considered extrinsic cohesion in the form of stabilizing selection (Collier, 1992, 1998, 2000). All the major transitions of Maynard Smith and Szathmary (1995, 1999) are associated with the emergence of a novel form of cohesion.
Functional integration and hierarchical structure are hallmarks of biological organization (e.g. Salthe, 1985, 1993; Wake and Roth, 1989 and references therein). A major component of functional integration is the interdependence of parts. Evolutionary changes in biological systems do not occur all at the same time; thus, when such changes occur, only part of the system changes. All changes, whether they are point mutations on a chromosome or alteration of part of a complex mating ritual, must integrate with the rest of the system, which has not changed, in such a way that viable organisms result. Hierarchical structure, from the genealogical relationships of individuals and species to the behavioral relationships of complex social systems and the trophic interactions among species in ecosystems, plays an important role in biological evolution. Salthe (1985, 1993) concluded that hierarchies provide stability, reinforce boundaries between system and surroundings, allow increasing amounts of complexity without losing organizational coherence, and provide a way in which causation and control can be tied together. He suggested that hierarchical structure can be decomposed into sets of "triads", comprising (1) upper level (causal or initiating), (2) lower level (control or boundary) elements impinging on (3) a focal level, from which emerges a particular form of structure and/or organization. Complex hierarchical systems are combinations of linked triadic units. Within a given hierarchical system, relatively random lower level effects are screened off by the cohesive properties of the higher level effects. In addition, the various levels in the hierarchy have diminishing effects on any given level in proportion to the remoteness of their interactions.
Salthe (1985, 1993; Eldredge and Salthe, 1984; Eldredge, 1985, 1986) has emphasized two major forms of hierarchically-organized biological structure. The ecological hierarchy is manifested by patterns of energy flow in ecosystems, and the genealogical hierarchy is manifested by patterns of ancestral relationships among organisms and species. Environmental and genealogical phenomena are good starting points for investigating hierarchical interactions because they are intimately connected in biology. Pre-biotic environmental conditions established the boundary conditions within which life could originate. Conversely, genealogical processes that characterize life are autonomous enough from environmental conditions to be capable of overrunning available resources and of changing the environmental conditions substantially (see section on natural selection below). The longer life exists on this planet, the more it shapes the environment of the planet. Today, much of the environment relevant to biological systems consists of products of genealogical processes (Maynard Smith, 1976; Brook and Wiley, 1986, 1988). The extent to which biological systems impose themselves on their environments, by creating their own niches (Brooks and Wiley, 1986, 1988; Odling-Schmee et al., 1996) or through evolutionary lag load (Maynard Smith, 1976) resulting from historical conservatism contributes greatly to the far from equilibrium “nature of the conditions” in which evolution takes place. Environmental effects under far from equilibrium conditions can lead to self-organization mediated by selection (Kauffman, 1993), so it is no surprise to discover that it is difficult to disentangle "environmental” effects from "genetic [genealogical]” effects in evolutionary studies.
The intimate relationship between the two hierarchies can be illustrated with a sports metaphor: The ecological hierarchy establishes the dimensions of the playing field, while the genealogical hierarchy establishes the rules of the game being played. In other words, biological systems obey rules of self-organization transmitted genealogically (historically) and played out within environmentally defined boundaries. To complicate matters, however, the self-organizing rules of the game by which living systems evolve can produce changes in the dimensions of the playing field. To extend the metaphor, the game may redefine the dimensions of the playing field, and may be subsequently constrained by those self-imposed changes. For example, the evolution of photosynthetic prokaryotes from anaerobic ancestors resulted in increased oxygen content in the atmosphere. This increase then altered the diversity and changed the distribution of anaerobic organisms, limiting them to relatively rare environments, and paving the way for the evolution of a new array of species. Now that we have a consistent view on the nature of the organism in a physical sense that also accounts for their Darwinian nature, we can discuss observed organismal diversity.
2 Re-Uniting The Nature Of The Organism With The Nature Of The Conditions: The Unified Theory And Selection Processes
The unified theory provides a coherent explanation for the inevitability of evolution, and for the origin and emergence of inherent orderliness in biological systems. It also relieves biologists of the necessity of invoking natural selection to explain every aspect of biological organization at every point in time. This is not to say that standard accounts of selection processes are irrelevant or outside the core of the unified theory (see also Collier, 1992, 1998). The unified theory is not only consistent with Darwinian views of evolution, it helps extend the Darwinian tradition by exposing and exorcising a specter looming in the background of Darwinian theory. The specter is that Darwinism and neo-Darwinism have never produced an explanation from first principles for the origin of their own central mechanism, natural selection. Darwin's original intuition about natural selection came by way of analogy with artificial selection informed in part by the writings of Malthus and Smith on human economic theory, and tied to the notion of the “nature of the organism” discussed at the beginning of this chapter. If natural selection is a real process, however, it must have some justification other than analogy with free-market capitalism. The following discussion results in a series of postulates chosen to emphasize the integration of the twin Darwinian principles of the nature of the organism and the nature of the conditions within the core of the unified theory.
As stated above, all the major transitions of Maynard Smith and Szathmary (1995, 1999) are associated with the emergence of a novel form of cohesion (e.g. chromosomes make genes cohesive, ontogeny makes cells cohesive, sex makes organisms cohesive). Each of these is also associated with a novel form of selection focused on that form of cohesion. Encoded information is also the carrier of the cohesive properties, so production of biological information involves simultaneous production of variation and constraints, ensuring that genealogy will be a combination of continuity and change.
Information Flow, the
Nature of the Organism and the Nature of the Conditions
Information systems consist of a source of signals, a channel through which the signals are transmitted, and a receiver to translate the portion of the signals that made it through the channel into information. Gatlin (1972) argued that the genetic system is the source, reproduction and ontogeny are the channel, and the environment is the receiver. Genetic possibilities thus become phenotypic signals as a result of reproduction and ontogeny, and become meaningful biological information as a result of causal interactions between the phenotype and the environment. The environment is not a receiver in a physical sense, because its only causal interaction with biological information is the possible elimination of some of it; it does not measure or interpret the information. Rather, at any given time the environment acts as interference in the channel. In energetic terms, environmental selection converts some biomass (enformation, ym) into "heat loss" (deS) through the elimination of relatively less fit organisms in a population. In informational terms, environmental selection acts as a "delete button" monitoring “incoming messages”, eliminating relatively less fit organisms (Brooks and McLennan, 1990, 2000; Brooks, 1991, 1992, 1994, 1997, 1998).
This raises the question of just what is “the receiver”. The answer lies in establishing that enformation is a material part of biological systems. If this is so, then it is possible for the system to be its own source and receiver (see also Rutledge et al., 1976; Csanyi, 1989). Current standard evolutionary theory stresses external causality as the ultimate source of orderliness and function in biological systems, so the receiver of genealogical information has been construed as part of the surroundings, i.e., localized in space. While it is true that biological systems are localized in space, they are also localized in time. Therefore, the receiver can be a "time" (Brooks and McLennan, 1990). The source is a genetic system at time t0, the channel is reproduction and ontogeny, and the receiver is the same genetic system at any given time t1…n; thus, the receiver is temporally distinct from the source (figure 2).
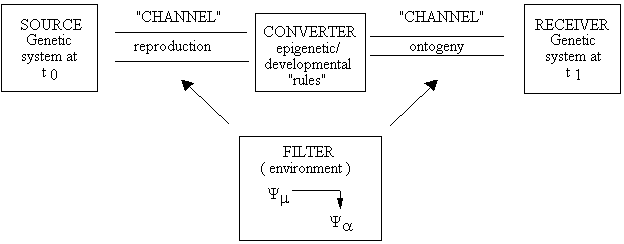
Figure 2. Biological information flow: (1) Source (gene pool at t0), encoder (reproduction), channel (ontogeny), noise (environment), receiver (genetic system at t1).
If the source precedes the receiver in time, it can produce the system that acts as receiver and that system can then become a source itself. A highly similar perspective has been used by information theorists designing self-correcting computer programs (Smith, 1998, 2000). This reinforces the biological analogy, because DNA has significant self-repair capabilities and sexual reproduction may enhance those capabilities (Bernstein et al., 1989). Finally, if the genetic system itself is the receiver, we can view environmental selection as contributing causally to evolutionary innovation through time, even if it is only eliminative at any given point in time. Such elimination can set the stage for future evolutionary creativity and innovation by altering the range of genotypes/phenotypes that survive the environmental filter at any point in time. Any such alterations would have the effect of changing historical genetic/developmental correlations in the system in the future, providing the opportunity for new evolutionary explorations.
Biological systems are physical information systems, a type of nonequilibrium thermodynamic system, open to exchanges of matter and energy but maintaining a closed information system internally which functions to reproduce the system, to perpetuate lineages through time. They are able to impose themselves and their functions on their surroundings, and thus are self-stabilizing and self-organizing. They produce organized complexity cheaply (diS is small compared to deS, and the portion of diS allocated for the information system is small; in part because a small number of chemical templates are used to generate many organisms), variably (because even chemical templates are subject to the statistical mechanical vagaries of the Second Law of Thermodynamics), and functionally (because organisms must exchange matter and energy both internally and with their surroundings in order to maintain themselves), but without regard for details of the surroundings (because the information system is embodied in relatively autonomous internal chemical production, diS, of the system). As the source and receiver of organized information, they can be the embodiment of the organizing principles for that information. Biological systems thus transmit information through, not to, their surroundings. This supports Darwin’s view that it is the [autonomous] nature of the organism that creates the necessary conditions for selection processes to occur.
Treating biological systems as physical information systems provides a causal basis for the origin of selection processes consistent with their well-documented causal consequences. Selection processes originate as a result of the necessity that biological systems obtain matter and energy from their surroundings coupled with the relative autonomy of their information systems, which permits production of organisms regardless of the details of their surroundings. Without the constraints provided by this autonomy, there would be no selection; at the same time, however, constraints provide systems with macroscopic properties that limit the ways in which and the extent to which the system will respond to selection. This means that biological systems should have their own macroscopic, or macroevolutionary, properties that are not reducible to microscopic dynamics. Some of the most important implications of re-emphasizing the nature of the organism in the context of thinking of evolution as an informational process involve the manner in which evolutionary theory views major events in the history of life (Maynard Smith and Szathmary, 1995, 1999). Below I will refer only to environmental and sexual selection as exemplars, but selection processes in this sense will emerge as causal mechanisms for any level of organization maintained by exchanges of matter and energy between system and surroundings in a form determined for the most part by information that resides relatively autonomously within the system itself.
2.1 The Origin Of Environmental Selection
The basis for the theory of natural selection was Darwin's observation that a major aspect of the “nature of the organism” in all species was that production of offspring overruns environmental resources necessary for their survival. This limitation leads to a "struggle for existence" in which only the "fittest" survive, fitness being defined by an individual's ability to compete for limited resources (its adaptedness), and measured by survival/reproductive success. Over the past century researchers have documented the intraspecific differences in survivability and reproduction vital to the concept of natural selection, and have set their explanations in an equilibrium context. A population is expected to reach a genetic (and hence informational) equilibrium with respect to its environment, and to remain there as long as the environment remains relatively constant. Subsequent to this, any environmental change creates a new equilibrium point, towards which the population moves (adapts). How can this equilibrium process be reconciled with the observation that biological systems and their evolution are far from equilibrium phenomena? The key to answering this question is understanding the two major conditions necessary for natural selection to exist in the first place.
First, the rules that lead to the production of organisms must be independent of, or at least highly insensitive to, fluctuations in the environment. Otherwise, the number of organisms produced would conform to a fluctuating equilibrium number determined by the availability of environmental resources, and we would not find different species exhibiting different reproductive rates in the same environments. Second, those rules must be characterized by a high degree of replication (redundancy), because if intraspecific competition is a driving force in natural selection, conspecific offspring must display overlapping requirements and abilities. Overall, then, if a large number of similar organisms are to be produced, the system's internal production rules must be relatively insensitive to environmental fluctuations (autonomy), and have a high degree of fidelity in replication (redundancy).
Biological systems are characterized by inherent production rules and a high degree of self-organization that is manifested, in part, by the maintenance of organized structure. The tendency towards increasing organization is not simply an effect of the environment. Naturally occurring populations of highly similar (redundant) organisms exist within boundaries defined both by genealogical history and by the environment. During relatively short time periods historical effects are essentially constant, and evolutionary dynamics can be explained solely by reference to environmental changes, corresponding to standard population biological treatments of natural selection. Over longer time periods, however, the stability and fate of a population will depend also on its history, which will constrain the ways in which and the extent to which the population can respond to environmental perturbations or changes. If this were not so, we would not have evidence that the environment can change so rapidly or in such a way that populations or species become extinct.
Eigen and Winkler (1981) discussed a thought experiment involving two chemotrophs living in the same environment and being identical in all ways except for different rates of replication. The chemotroph with the higher replication rate will always outcompete the one with the slower rate. This thought experiment was designed to show natural selection as a primitive concept, but it works only if the different replication rates of the two chemotrophs in the same environment are assumed to be intrinsic properties of the chemotrophs; that is, that the two chemotrophs are self-determining rather than environmentally-conforming with respect to their replication rates. Thus, natural selection seems to require some degree of autonomy. The thought experiment also shows that the fitness of the two chemotrophs relative to each other is not an intrinsic property of either, but rather results from the interaction between them and their environments. The two chemotrophs might each exhibit different replication rates in different environments (possibly either of the chemotrophs might have the slower replication rate in one environment and the faster one in a different environment), and it is also possible that each of the chemotrophs might come into contact with yet other chemotrophs in yet other environments in which the same kinds of arguments hold true. Thus, Darwin was correct in asserting that the nature of the organism is more important than the nature of the conditions, and that this holds the key to understanding how important is environmental selection in evolution (see also Maynard Smith and Szathmary , 1995, 1999 for a similar argument). Or,
Postulate 1: In order for environmental selection
to be a real process, there must be rules governing the production of organisms
that are largely insensitive to environmental resources, in addition to a
causal link between the system and its surroundings. If this is true, however,
those rules may affect the ways in which, and the extent to which, populations
respond to selection.
2.1.1 Collier's Paradox
We expect
macroscopic information (I), or organization, to increase over time in evolving
systems (for exceptions see Smith, 1998); therefore, we expect to see an
increase in the value of the function Q, the macroscopic order (Landsberg,
1984b) of the system:
Q
= 1 - (Hobs/Hmax) = Id/Hmax
If the environment is the primary source of macroscopic ordering for biological systems, then the percentage of biological information that is shared between system and environment should be high. This presents a paradoxical view of natural selection because, if the amount of this mutual information is high, the "fit" between biological systems and their environments is almost perfect; therefore, evolutionary change will be negligible or stochastic with respect to the environment. This paradox can be resolved by postulating that the amount of information shared between biological systems and their environments is low (Collier, 1986). If this is true, the number of organisms requiring a particular environmental resource (or set of resources) may exceed the availability of that resource, and natural selection will emerge as an evolutionary process. Thus, in order for natural selection to be an important evolutionary force, it must operate under conditions established by genealogically driven self-organization which, in turn, produce constraints on the way in which and the extent to which populations can respond to natural selection. In other words, natural selection co-emerged with genetically autonomous living systems, and has thus always been (and always will be) part of the process (Maynard Smith and Szathmary, 1995, 1999).
The unified theory thus explains why there is natural selection at all, and at the same time it suggests limits on the effects of natural selection because it operates in a context defined by genealogical phenomena, which themselves are inherently evolutionary. From the externalist perspective, the environment is always decaying with respect to the population average fitness, and inherent constraints limit the rate at which, the ways in which, and the extent to which the population can respond. This leads to what Maynard Smith (1976) termed "lag load", what Ulanowicz (1986, 1997) termed "ascendancy", and what others have called “phylogenetic constraints” or “phylogenetic inertia” (see also Brooks, 1997, 1998; Brooks and McLennan, 2000, in press). The environment is always deteriorating with respect to the most fit genotype, and this creates an environmental "pull" to which genealogical systems respond in a manner that leaves room for the genealogical system to "experiment".
Genetic systems have three kinds of relationship with their environments: (1) complementary, in which case genealogical and environmental dynamics will be in phase and the inherent properties of the genetic system will be promoted by the environment; (2) neutral, in which case the genealogical system and the environment may change out of phase with each other, with no net adverse impact on the genetic system (I will discuss a possible example of this later); and (3) antagonistic, in which case the genetic system will be restricted by the environment. Relationship (3) will rarely be observed in macroevolutionary patterns, because the portions of the genealogical system that have been "selected out" leave little macroevolutionary evidence of their existence. Cases (1) and (2) may not be distinguishable in practice, except possibly by the emergence of convergent adaptations (Brooks and McLennan, 1991, in press). Selection effects are expected to be microscopically ubiquitous, tending to optimize populations locally in terms of the immediately environment, to the extent possible given the developmental, reproductive, behavioral, and phylogenetic cohesion of the species. Such effects can contribute to macroscopic behavior to the extent that they limit the range of variation that participates in reproduction, eliminating some otherwise functional genetic variants. Environmental selection can thus act as a form of external cohesion on the species (Collier, 1998, 2000; Collier and Hooker, 1999). Rates of environmental change tend to be higher than rates of speciation, so environmental selection should contribute far more to microscopic than macroscopic phenomena. Thus, the resolution of Collier's Paradox is
Postulate 2: The effectiveness of environmental
selection will be inversely proportional to the amount of mutual information
between the genealogical system and the environment. If genealogical changes do
not occur in response to, and/or if they occur more slowly than, environmental
changes, the genealogical system will lag behind the environment (Maynard
Smith, 1976), and will always be "trying to catch up" in the
"Red Queen" sense. The extent to which this is true is the extent to
which natural selection will always be potentially an evolutionary force. That
is, there will always be ecological ascendancy or "unoccupied niche
space", allowing for experimentation by the genetic system, but only so
long as genealogical changes do not
track environmental changes closely.
2.1.2 The Origin of Sexual Selection
Sexual selection emerges from increased mutual information between members of the genealogical hierarchy; thus, it is quite different from natural selection, just as Darwin thought. The degree to which sexual selection is effective is directly proportional to the degree of mutual information, and it is genealogical processes that are responsible for shared information (genealogical and phylogenetic constraints), so sexual selection should be an evolutionarily cohesive process. Thus,
Postulate 3: In order for sexual selection to be a
real process, males and females must be able to communicate with each other in
such a way that sexual preferences influence the flow of genetic information
from one generation to another. Thus,
sexual selection should be a cohesive evolutionary process.
2.1.3 The Difference between Environmental and Sexual Selection
Figure 3 portrays the differences between natural and sexual selection using the formalism summarized in figure 2.

Figure 3. Environmental and sexual selection in biological information flow.
Sexual selection operates at the source of information transmission, which is reproduction, and tends to increase the efficiency of encoding information in such a way that members of the next generation express heritable mate preferences. Natural selection, by contrast, operates at the endpoints of information transmission, which are the products of reproduction, and tends to decrease the capacity of the channel through which information is transmitted, so that a smaller fraction of all possible bits of information actually get through to the next generation. Or,
Postulate 4: Environmental selection emerges as a result of a lack of sensitivity (low mutual information) between the genetic system and the environment. Sexual selection, by contrast, emerges as a result of increased sensitivity (high mutual information) between members of the genetic system. Environmental selection reduces variance by restricting the capacity of the channel through which information can be transmitted successfully. Sexual selection, by contrast, reduces variance by restricting the range of variants encoded at the (reproductive) source. Thus, for environmental selection, the probability of survival is the result of interfering events, whereas for sexual selection the probability of survival is the result of non-response to interfering events.
The unified theory postulates that genealogical systems always "mean something" or "talk to" themselves (e.g., Csanyi, 1989; Brooks and McLennan, 1990, 1997; McLennan, 1997). For an organism to "mean something to itself", it must develop to the extent of becoming reproductively competent. Because it is also organized by the environment in which it resides, organisms also "mean something" or "talk to" their surroundings, and the evolutionary measure of what an organism means to its environment is fitness (Brandon, 1990; see also Collier, 1992). Metaphorically speaking, we can see that evolution is fundamentally a Darwinian process because whenever there is a conflict between what an organism means to itself (which includes sexual selection) and what it means to its environment (natural selection), self-meaning always takes precedence, with a resulting decrease in fitness of the individual. Once again, it is the primacy of the “nature of the organism” over the “nature of the conditions” that make natural selection a powerful force, but only within the constraints provided by the nature of the organism.
Natural and sexual selection may exhibit three different types of relationship with respect to each other. If antagonistic, sexual selection might tend to promote evolutionary trends that are maladaptive with respect to environmental selection; this corresponds to "runaway sexual selection" scenarios. If complementary, sexual selection will look like a subset of natural selection; this corresponds to "truth in advertising" scenarios. Finally, if neutral, each type of selection influence will tend to modify different portions of the genealogical system (different characteristics) at the same time. Therefore,
Postulate 5: Environmental and sexual selection
arise in different ways, and affect different parts of the information flow
system, so it is possible for them to be in conflict (one possibility in
Fisherian runaway sexual selection theory, in which sexual selection could lead
to efficiently encoded information in such a way that less and less of it could
get through the channel permitted by the environment), or to complement each
other ("truth in advertising" views of sexual selection, in which
sexual selection could lead to efficiently encoded information in such a way
that it flows through the channel with reduced distortion).
3 Conclusions: The Nature of Evolution
In adaptive radiation and in every
part of the whole, wonderful history of life, all
the modes and all the factors of
evolution are inextricably interwoven.
The total
process cannot be made simple, but
it can be analyzed in part. It is not understood
in all its appalling intricacy,
but some understanding is in our grasp, and we may
trust our own powers to obtain
more. Simpson 1944
The emerging unified theory treats biological evolution metaphorically as a microcosm of cosmological evolution. The expansion of the matter in the universe increases the actual entropy of the universe, in accordance with the second law. Gravitational effects and other forms of symmetry-breaking that emerge from that initial expansion, however, slow the disorganization of the universe, so the actual increase in entropy is less than maximal. On the energetic side, the unified theory treats the energy/entropy relationship as highly relative. From the perspective of autotrophic biological systems solar heat and light energy is free energy. From the perspective of the sun, however, that heat and light is entropy, because it represents relatively low-grade energy lost from the sun as a result of thermonuclear reactions. Some solar energy is picked up by autotrophs on this planet and converted into two forms of entropy, heat lost from the plant and organic structure. I believe this metaphorical perspective is similar to Boltzmann's, who wrote that he considered the 19th century to be Darwin’s Century, and disagreed with Darwin only to the extent that he thought we would eventually find that evolution was not so much a struggle for survival as a struggle for entropy [production] (Brush, 1983), including the entropy of solar reactions, some of which can serve as free energy for biological systems. The biological phase space comprises the inheritance systems (Maynard Smith and Szathmary, 1995, 1999), and it expands over time as mutations add new dimensions to it. This increases the maximum possible entropy of the system, making it a dynamic environment for evolving systems. However, the effects of common ancestry and of the environment on mate finding, reproduction, and development, provide constraints on the increases in the entropy (diversity) of the biological information system.
The storage and transmission of information from one generation to the next (biological production that functions solely within the genealogical hierarchy) uses a tiny fraction of the total energy budget of an organism, so it is not an energy-limited (Brooks and Wiley, 1988) or metabolism-limited (Maynard Smith and Szathmary, 1995, 1999) process. The rest of biological production, which functions in the ecological hierarchy, can be viewed as an emergent property of the genealogical hierarchy. Biomass is produced by genealogical rules but functions in the ecological realm; only genetic information functions solely within the genealogical hierarchy, and it does not cost very much energetically. The tiny portion of the total energy budget of biological systems invested in the storage and transmission of genetic information, however, is at least as important as the rest of the energy budget in explaining biological diversity. We believe that what controls the origins of evolutionary diversity takes up such a negligible portion of the total energy budget of biological systems that it has been generally overlooked. Many genealogical processes appear negentropic from the perspective of the environment, because so much of the entropy production (i.e., so large a proportion of the total energy budget) of genealogical systems is returned to the surroundings; hence, the surroundings tend to decay more rapidly than the systems and this could be (and has been) misinterpreted as negentropic behavior on the part of the biological system. Organisms do not just degrade their immediate environments; they can serve as environmental sources of energy for other organisms; in fact, the largest portion of the environment for organisms is other organisms. Organism functions that increase the amount of energy, and the amount of time energy from abiotic sources remains in biological systems, thus represent evolutionary mechanisms by which the rate of environmental degradation can be slowed. This is done this by organisms sequestering entropy production for their own use, a purely selfish behavior that nonetheless benefits others (Matsuno, 1989, 1995, 1996, 1998, 2000).
Finally, I believe the emerging unified theory of evolution is characterized by the recognition that there is no single objective level of organization, spatial scale, or temporal scale that has causal primacy in biological evolution. Neither intrinsic nor extrinsic processes are primary, much less exclusive, over all spatial or temporal scales, for all types of biological interactions, or over all groups of organisms. A variety of processes operate on many different scales, and all contribute to evolution; however, they do not all play equally important roles at all levels, at all times, or for all groups. Therefore, the macroscopic manifestations of evolutionary principles will differ depending on the group of organisms and the window of observation. It is a theory that returns us to the panoramic view of life first given us by Darwin.
4 Acknowledgements
My perspective on evolution has itself evolved over the past 20 years, due in no small measure to the willingness of many colleagues to engage me on various topics. They have helped me refine my arguments for supporting certain views, but more importantly, they have helped me understand the limitations and incorrectness of many of my views. I have no hope of ever eliminating all my misunderstandings, but trying to do so has been an exciting, as well as humbling and enlightening experience. Particular thanks to Mishtu Banerjee, Douglas Causey, John Collier, Cyril Finnegan, Rob Geesink, Hironori Hirata, Cliff Hooker, Deborah McLennan, Koichiro Matsuno, Brian Maurer, Jack Maze, Stan Salthe, James D. Smith, Jonathan Smith, Eors Szathmary, Robert Ulanowicz, Edward Wiley and Rino Zandee. Particular thanks to John Collier for help with this manuscript. Support for this research was provided by operating grant A7696 from the Natural Sciences and Engineering Council (NSERC) of Canada.
References
Bernstein, H., F.A. Hopf, and R.E. Michod. 1988. Is meiotic recombination an adaptation for
repairing DNA, producing genetic variation, or both? In The Evolution of Sex, ed.
Michod, R.E. and B.R. Levin. p 139-160. Sunderland Mass.: Sinauer Assoc.
Boltzmann, L. von. 1877. Uber die Beziehung eines allgemeine mechanischen Satzes zum
zweiten Haupsatzes der Warmtheorie. Sitzungsber. Akad. Wiss. Wien, Math.-Naturwiss.
Kl. 75: 67-73.
Bowler, P.J. 1983. The Eclipse of Darwinism. Baltimore: Johns Hopkins Univ. Press.
Brillouin, L. 1962. Science and Information Theory. 2nd ed. New York: Academic Press.
Brooks, D.R. 1988. Scaling effects in historical biogeography: A new view of Space, Time and
Form. Syst. Zool. 37: 237-244.
Brooks, D.R. 1990. The unified theory, macroevolution, and historical ecology. In The Plant
Diversity of Malesia, ed. P. Baas et al. pp. 379-386. Amsterdam: Kluwer.
Brooks, D.R. 1992. Incorporating origins into evolutionary
theory. In Understanding Origin:
Contemporary Ideas on the Genesis of Life, Mind and Society, ed. F. Varela and J. P.
Dupuy. pp. 191-212. Amsterdam: Reidel/Kluwer Associates.
Brooks, D.R. 1994. Entropy, information and evolving biological systems. Theor. Hist. Scient. 4:
31-49.
Brooks, D.R. 1997. Biological evolution as a microcosm of cosmological evolution. Bridges 4:
9-35.
Brooks, D.R. 1998. The unified theory of evolution and
selection processes. In Evolutionary
Systems:
Biological and Epistemological Perspectives on Selection and Self-
Organization, ed. G. van de Vijver, S.N. Salthe, and M. Delpos, 113-128. Dordrecht:
Kluwer Academic Publ.
Brooks, D.R. 2000. The nature of the organism: life takes on
a life of its own. Proc. N.Y. Acad.
Sci. 901: 257-265.
Brooks, D.R. 2001. Diversity, organismal level. In Encyclopedia of Biodiversity. Vol. 2: 191-
207. Ed. S. Levins. New York: Academic Press.
Brooks, D.R., J. Collier, B.A. Maurer, J.D.H. Smith, and E.O. Wiley. 1989. Entropy and
information in evolving biological systems. Biol. Philos. 4: 407-432.
Brooks, D.R., D.D. Cumming and P.H. LeBlond. 1988. Dollo's Law and the second law of
thermodynamics: analogy or
extension? In Entropy, Information and
Evolution: New
Perspectives on Physical and Biological Evolution, pp. 189-224, ed. B. Weber, D.J.
Depew, and J.D. Smith. MIT Press, Cambridge.
Brooks, D.R., P.H. LeBlond and D.D. Cumming. 1984. Information and entropy in a simple
evolution model. J. Theor. Biol. 109: 77-93.
Brooks, D.R. and D.A. McLennan. 1990. Searching for a general theory of biological evolution.
J. Ideas 1: 35-46.
Brooks, D.R. and D.A. McLennan. 1991. Phylogeny, Ecology and Behavior: A Research
Program in Comparative Biology. Univ. Chicago Press, Chicago.
Brooks, D.R. and D.A. McLennan. 1997. Biological signals as
material phenomena. Rev.
pensee
d’aujord d’hui 25: 118-127.
Brooks, D.R. and D.A. McLennan. 2000. The nature of the organism and the emergence of
selection processes and biological signals. In Semiotics, Evolution, Energy. E. Taborsky,
ed. pp. 185-218. Aachen, Germany: Shaker Verlag.
Brooks, D.R. and D.A. McLennan. in
press. The Nature of Diversity: A Voyage
Through Space
and Time. Chicago: Univ. Chicago Press.
Brooks, D.R. and E.O. Wiley. 1986. Evolution as Entropy: Toward a Unified Theory of Biology.
1st ed. Univ. Chicago Press, Chicago.
Brooks, D.R. and E.O. Wiley. 1988. Evolution as Entropy: Toward a Unified Theory of Biology.
2nd ed. Univ. Chicago Press, Chicago.
Brush, S.G. 1983. Statistical
Physics and the Atomic Theory of Matter, from Boyle and Newton
to Landau and Onsager. Princeton: Princeton Univ. Press.
Collier, J. 1986. Entropy in evolution. Biol. Philos. 1: 5-24.
Collier, J. 1988. The dynamics of biological order. In Entropy, Information and Evolution: New
Perspectives on Physical and Biological Evolution, pp. 227-242, ed. B. Weber, D.J.
Depew, and J. D. Smith. MIT Press, Cambridge.
Collier, J. 1990. Two faces of Maxwell's demon reveal the
nature of irreversibility. Stud. Hist.
Philos. Sci.
Collier, J. 1992. Incororporating adaption into the unified theory.
http://www.newcastle.edu.au/department/pl/Staff/JohnCollier/papers/iaut.pdf
Collier, J. 1998. Information increase in biological
systems: How does adaptation fit? In
Evolutionary
Systems: Biological and Epistemological Perspectives on Selection and
Self-Organization, ed. G. van de Vijver, S.N. Salthe, and M. Delpos, 129-140. Dordrecht:
Kluwer Academic Publ.
Collier, J. 2000. The dynamical basis of information and the
origins of semiosis. In Semiotics,
Evolution, Energy. E. Taborsky, ed. P. 111-138. Aachen, Germany: Shaker Verlag.
Collier, J. and C. Hooker. 1999. Complexly organised dynamical systems. Open Syst. Info. Dyn.
6: 241-302. http://www.newcastle.edu.au/department/pl/compsys/publications/Cods.pdf
Csanyi, V. 1989. Evolutionary Systems and Society: A General Theory. Durham, N.C.: Duke
Univ. Press.
Darwin, C. 1859. Origin of Species. London: J. Murray.
Darwin, C. 1872. The Origin of Species. London: John Murray. 6th edition.
Demetrius, L. 1992. The thermodynamics of evolution. Physica A 189: 417-436.
Depew, D. and B. Weber. 1995. Darwinism Evolving. Cambridge, Mass.: Bradford Books of
MIT Press.
Eigen, M. and R. Winkler. 1981. Laws of the Game: How the Principles of Nature Govern
Chance. New York: A.A. Knopf.
Eldredge, N. 1985. Unfinished Synthesis. New York: Columbia Univ. Press.
Eldredge, N. 1986. Information, economics and evolution. Ann. Rev. Ecol. Syst. 17: 351-369.
Eldredge, N. and S. N. Salthe. 1984. Hierarchy and
evolution. In Oxford Surveys in
Evolutionary
Biology, ed. R. Dawkins and M. Ridley 1: 182-206.
Frautschi, S. 1982. Entropy in an expanding universe. Science 217: 593-599.
Frautschi, S. 1988. Entropy in an expanding universe. in Entropy, Information and Evolution:
New Perspectives on Physical and Biological Evolution, pp. 11-22, ed. B. Weber, D.J.
Depew, and J. D. Smith. Cambridge: MIT Press.
Gatlin, L.L. 1972. Information Theory and the Living System. New York: Columbia Univ. Press.
Gladyshev, G.P. 1996. Thermodynamic direction of biological evolution: model and reality.
Izvestiya akademii nauk seriya biologicheskaya 4: 389-397.
Goodwin, B. 1995. How the Leopard Changed its Spots. London: Phoenix Giants.
Gould, S. J. and R. C. Lewontin. 1979. The spandrels of San Marco and the Panglossian
paradigm: A critique of the adaptationist programme. Proc. R. Soc. London B205: 581-
598.
Hirata, H. 1993. Information of organization in ecological systems: Nutrient > Energy > Carbon.
J. theor. Biol. 162: 187-194.
Kampis, G. 1991. Self-modifying
systems in Biology and Cognitive Science: A New Framework
for Dynamics, Information and Complexity. Oxford: Pergamon.
Kampis, G. 1998.Evolution as its own cause and effect. In Evolutionary Systems: Biological and
Epistemological Perspectives on Selection and Self- Organization, ed. G. van de Vijver,
S.N. Salthe, and M. Delpos, 255-265. Dordrecht: Kluwer Academic Publ.
Kauffman, S. A. 1993. The Origins of Order: Self-organization and Selection in Evolution. New
York: Oxford Univ. Press.
Kjellstrom, G. 1996. Evolution as a statistical optimization algorithm. Evol.Theory 11:
105-117.
Landsberg, P.T. 1984a. Is equilibrium always an entropy maximum? J. Stat. Physics 35: 159-
169.
Landsberg, P.T. 1984b. Can entropy and "order" increase together? Physics Lett. 102A: 171-173.
Layzer, D. 1975. The arrow of time. Sci. Amer. 233: 56-69.
Layzer, D. 1978. A macroscopic approach to population genetics. J. Theor. Biol. 73: 769-788.
Layzer, D. 1980. Genetic variation and progressive evolution. Amer. Nat. 115: 809-826.
Lemke, N. 1998. The phenotype space approach to prey predator coevolution. Theory in
Biosciences 117: 321-333.
Lotka, A.J. 1913. Evolution from the standpoint of physics, the principle of the persistence of
stable forms. Sci. Amer. suppl. 75: 345-6, 354, 379.
Lotka, A.J. 1925. Elements of Physical Biology. Williams and Wilkins, Baltimore.
McLennan, D.A. 1997. Signals. Semiotics Review of Books 8: 1-3.
Matsuno, K. 1989. Protobiology: Physical Basis of Biology. Boca Raton, Florida: CRC Press.
Matsuno, K. 1995. Consumer power as the major evolutionary force. J. theor. Biol.. 173: 137-
145.
Matsuno, K. 1996. How many trophic levels are there? J. Theor. Biol. 180:105-109.
Matsuno, K. 1998. Competence of natural languages for
describing the physical origin of life. In
Evolutionary Systems:
Biological and Epistemological Perspectives on Selection and
Self- Organization, ed. G. van de Vijver, S.N. Salthe, and M. Delpos, 295-306.
Dordrecht: Kluwer Academic Publ.
Matsuno, K. 2000. Material contextualization in time. In Semiotics, Evolution, Energy. E.
Taborsky, ed. pp. 219-230. Aachen, Germany: Shaker Verlag.
Maurer, B.A. and D.R. Brooks. 1991. Energy flow and entropy production in biological systems.
J. Ideas. 2: 48-53.
Maynard Smith, J. 1970. Time in the evolutionary process. Studium Generale 23: 266-272.
Maynard Smith, J. 1976. What determines the rate of evolution? Amer. Nat.. 110: 331-338.
Maynard Smith, J. and E. Szathmary. 1995. The Major Transitions in Evolution. Oxford: W.H.
Freeman Spektrum.
Maynard Smith, J. and E. Szathmary. 1999. The Origins of Life. Oxford: Oxford Univ. Press.
Niklas, K. J. 1999. Evolutionary walks through a land plant morphospace. J. Exper. Bot. 50: 39-
52.
Odling-Schmee, F.J., K. N. Laland, and M.W. Feldman. 1996. Niche construction. Amer. Nat.
86: 309-326.
Prigogine, I. 1980. From Being to Becoming. W. H. Freeman, San Francisco.
Prigogine, I. and J.M. Wiame. 1946. Biologie et thermodynamique des phénomènes
irréversìbles.Experientia 2: 451-453.
Raff, R.A. 1996. The Shape of Life: Genes, Development, and the Evolution of Animal Form.
Chicago: Univ. Chicago Press.
Rose, M.R. and G.V. Lauder (eds.). 1996. Adaptation. NY: Acadmic Press.
Rutledge, R.W., B.L. Basorre, and
R.J. Mulholland. 1976. Ecological stability: an information
theory
viewpoint. J. theor. Biol.
57:355-371.
Salthe, S.N. 1985. Evolving Hierarchical Systems: Their Structure and Representation.
Columbia Univ. Press, New York.
Salthe, S.N. 1993. Development and Evolution: Complexity and Change in Biology. MIT Press,
Boston.
Schanck, J.C. and W.C. Wimsatt. 1988. Generative retrenchment and evolution. In PSA 86, vol.
2, ed. A. Fine and P.K. Machamer, p. 33-60. Philosophy of Science Association, East
Lansing, Michigan.
Shpak, M. and G.P. Wagner. 2000. Asymmetry of configuration space induced by unequal
crossover: implications for a mathematical theory of evolutionary innovation. Artif. Life
6: 25-43.
Simpson, G.G. 1944. Tempo and Mode in Evolution. New York: Columbia Univ. Press.
Smith, J.D.H. 1988. A class of mathematical models for evolution and hierarchical information
theory. Inst. Math. Appl. Preprint Series 396: 1-13.
Smith, J.D.H. 1998. Canonical ensembles, competing species,
and the arrow of time. In
Evolutionary Systems:
Biological and Epistemological Perspectives on Selection and
Self-Organization, ed. G. van de Vijver, S.N. Salthe, and M. Delpos, 141-154. Dordrecht:
Kluwer Academic Publ.
Smith. J. D.L. 2000. On the evolution of semiotic capacity. In Semiotics, Evolution, Energy. E.
Taborsky, ed. P. 283-309. Aachen,
Germany: Shaker Verlag.
Taborsky, E. ed. 2000. Semiotics, Evolution, Energy. Aachen, Germany: Shaker Verlag.
Ulanowicz, R.E. 1986. Growth and Development: Ecosystems Phenomenology. New York:
Springer-Verlag.
Ulanowics, R.E. 1997. Ecology: The Ascendent Perspective. New York: Columbia University
Press.
Van de Vijver, G., S.N. Salthe, and M. Delpos. eds. 1998.
Evolutionary Systems: Biological and
Epistemological Perspectives on Selection and Self-Organization. Dordrecht: Kluwer
Academic Publ.
Wake, D.B. and G. Roth. eds. 1989. Complex Organismal Functions: Integration and Evolution
in Vertebrates. Dahlem Workshop. New York: Wiley.
Weber, B.H. and D.J. Depew. 2000. The modern evolutionary synthesis and complex systems
dynamics: prospects for a new synthesis. In Semiotics, Evolution, Energy. E. Taborsky,
ed. P. 263-282. Aachen, Germany: Shaker Verlag.
Weber, B., D. Depew, and J. Smith. 1988. Entropy, Information and Evolution. Cambridge MA:
MIT Press.
Wicken, J.S. 1987. Evolution,
Thermodynamics and Information: Extending the Darwinian
Paradigm. Oxford Univ. Press: Oxford.
Wiley, E.O. 1981. Phylogenetics: The Theory and Practice of Phylogenetic Systematics. New
York: Wiley-Intersci.
Wiley, E.O. and D.R. Brooks. 1982. A nonequilibrium approach to evolution. Syst. Zool. 31: 1-
24.
Zotin, A.I. and R.S. Zotina. 1978. Experimental basis for qualitative phenomenological theory of
development. in Thermodynamics of Biological Processes, pp. 61-84, ed. I. Lamprecht
and A.I. Zotin. deGruyter, Berlin.