The Interrelations between Genotype/Phenotype/Environment:
A Semiotic Contribution to
the Evo:Devo Debate
Eugenio
Andrade
Professor
Department of Biology
Universidad
Nacional de Colombia
Phone: 57-1-3165000 Ext. 11313
Email: leandradep@unal.edu.co
Mailing address: A.A. 330413.
© This paper is not for reproduction without the permission of the author.
Abstract
Seemingly opposite views of evolution and development that conflict over the role assigned to either or structure/function, genes/environment, random/directed variations, innate/acquired characteristics, instructive/selective information, self-organization/natural selection, are still hotly debated. This conflict results from the self-referential loop present in the ontic (internal/external) and epistemic (individual-local/population-global) cuts, that together contribute to integrate developmental and evolutionary theories. It is assumed that development and evolution are non-programmed open ended processes of information increase. Organisms are modeled as Evolving Developing Agents that codify environmental information and possesses a general transformative tendency to reach a functional compromise between: a) Increments of phenotype’s uniqueness (stability and specificity), and b) Anticipation to environmental changes. Accordingly, changes in mutual information content between the Phenotype/Environment drag subsequent changes in mutual information content between Genotype/Phenotype and Genotype/Environment at two parallel and interwoven scales: individual life cycle (ontogeny) and species time (phylogeny) respectively. Developmental terminal additions along with the minimization of developmental steps must be positively selected. This approach shows how the six Peircian space/time/function relations described by Taborsky provide a conceptual framework that favors this unifying view.
Keywords: Genotype (G), Phenotype (P), Environment (E),
development, evolution, information, self-reference, Evolving Developing Agents
(EDA), semiosis.
1. Introduction
The Lamarckian internal/external cut:
The problem about the manner in which the relations
between organisms and environment influence evolutionary change goes back to
the birth of modern biology with Lamarck’s distinction between internal and
external realms. A divide made on the assumption that a spontaneous rupture of
homogeneous symmetry was associated with the generation of life and the environment.
This distinction was a sine qua non
condition for the foundation of biology as a natural science by providing it
with an object with intrinsic lawful properties. This is equivalent to the
ontological cut (Atmanspacher 2002, Taborsky 2002, 2004), for it defined a
biological realm as ontologically distinct from the physico-chemical inorganic
world. Organisms were, then, conceived as sentient beings with inner impulses
and organized structure that enabled them to respond to external environmental
influences. The Inner Sentiment or
capacity to act in a coordinate way was directly dependent on organization
(Burkhardt 1995). The increasing differentiation was associated to an ever
higher sophistication of the systems of inner coordination so that the activity
of the organisms as a coherent and cohesive whole was preserved (Lamarck 1803).
The transformations of living forms were perceived as embryological processes
for which nature employed intrinsic and extrinsic factors. The former is the inner
sentiment that by means of heat (vital force) tends to make organization more complex. The latter
corresponds to the conditions of life or the environment that exert a direct
action on the structure and heredity of living beings (Jacob 1982). The Lamarckian ontological cut is between
organism’s inner impulse and their environment that shapes their external
appearance through a process of irreversible transformations that makes
time the essential dimension of organic reality as the world of possibilities in
the here now (present tense) brings forth actual realizations (perfect
tense). The upper quadrants in figure 1, illustrates
individual development as a transition from II (internal individual
potentiality) to I (external individual completion).
In agreement with an
embryological perspective,
For the embryo is the animal in its less
modified state; and in so far it reveals the structure of its progenitor. …
Thus, community in embryonic structure reveals community of descent. … As the
embryonic state of each species and group of species partially shows us the
structure of their less modified ancient progenitors, we can clearly see why
ancient and extinct forms of life should resemble the embryos of their
descendants, our own existing species.
Following Richards (1992)
These
two divisions, Ontogeny, (embryonic development of the individual) or the
history of the germ, Phylogeny, or the history of the tribe, are intimately
connected, and the one cannot be understood without the other ... Ontogeny is a
recapitulation of Phylogeny; … the series of forms through which the individual
organism passes during its progress from egg cell to its fully developed state,
is a brief, compressed reproduction of the long series of forms through which
the animal ancestors of that organism ... have passed from the earliest periods
of so called organic creation down to present time. (Haeckel 1879).
Although the processes of the transformation of individuals and species undoubtedly share common properties they ought to be differentiated because an approach founded solely on the internal/external distinction confuses individual and population levels. A new cut conceives a more subtle distinction within the ontic realm.
(Figure 1)
The Neodarwinian individual-local/population-global cut:
This is a distinction for which
If the laws of nature are results of evolution, this evolution must proceed according to some principle, and this principle will itself be of the nature of a law. But it must be such a law that it can evolve or develop itself. (…) Evidently it must be a tendency toward generalization, - a generalizing tendency. But any fundamental universal tendency ought to manifest itself in nature. Where shall we look for it? (…) But we must search for this generalizing tendency rather in such departments of nature where we find plasticity and evolution still at work. The most plastic of all things is the human mind, and next after that comes the organic world, the world of protoplasm. Now the generalizing tendency is the great law of mind, the law of association, the law of habit taking. We also find in all active protoplasm a tendency to take habits. Hence I was led to the hypothesis that the laws of the universe have been formed under a universal tendency of all things toward generalization and habit taking. (Peirce, 1898. C.P. 7.515).
Genetics rests on a foundation given by the existence
of a Weissman’s barrier, according to which the phenotype was determined by the
prescriptions encoded in G so that the modifications of P by the action of E cannot
affect G, a way of reasoning inspired the central dogma of molecular biology in
the nineteen fifties. Thus, the G/P duality became the new version of the
internal/external duality where the self was identified with the gene as an
individual and as a population. Accordingly Neodarwinians focused on the shift
in the genetic composition of the population by the effects of natural
selection.
In consequence, individual organisms were stripped of all autonomy and came to be imagined as determined by two opposing causes, an efficient cause operating upwards from the lower levels (G) and a final cause operating downwards from the higher levels (E). Neodarwinism considers that the environment poses challenges that are to be solved by the organisms, in a process of adaptation in which it is a priori assumed that among the random variants, very few preexist in a population that possesses an adequate fit to the environment. This is depicted as a movement from quadrant IV to III in figure 1, which is mediated by the ever going action of natural selection in the progressive time.
Natural Selection is daily and hourly scrutinizing,
throughout the world, every variation, even the slightest; rejecting
that which is bad, preserving and adding up all that is good; silently
and insensibly working, whenever and wherever opportunity offers, at the
improvement of each organic being in relation to its organic and inorganic
conditions of life. (
The statistical mechanical conception provides statistical predictability under specific boundary conditions by means of the identification of fundamental units and interaction rules among them. But when the intrinsic dynamics and activity of the individual organisms occurring in the two upper quadrants (figure 1) is neglected, the formulation of an integrated evolutionary and developmental perspective is hampered. So long as individuals are not thought as a real information processing agents, “randomness” and “selection” are exclusively confined to properties of the genetic and environmental factors respectively. With the neglect of individual autonomy, intention and teleological interpretations were drastically eliminated in the workings of nature and in consequence embryology was done away, since it was conceived as preprogrammed teleological dead ended processes.
The Epigenetic View and
Developmental Systems Theory (DST):
There is an ongoing debate about the relations between ontogeny and phylogeny; each one at its turn has been postulated to be the efficient cause of the other. The internal domain was the level where coherence was achieved and individual evolutionary variations turn up, but it was easier to assume their random production instead of asking how developmental processes result in individual variants. Mainstream biologists prefer to affirm that embryos develop according to a genetic program rather than being subject to other influences. In the last decades the discovery of Hox genes revived an interest in development among geneticists that manage to subsume development to a higher hierarchy of controlling genes and so placed even less weight to factors other than genes. But to what extent, is ontogeny a deterministic process guided by the information contained in the genes? To what extent is evolution exclusively directed by natural selection in a specific environment? By contrast, the epigenetic perspective considers ontogeny as a process in which some additional information from the environment is incorporated in order to produce the form. Then, the relationship between G and P is sensitive to circumstantial fluctuations under concrete local restrictions. Defenders of the epigenetic view consider genetics as a new edition of preformism according to which no real epigenesis takes place since all formative factors preexists in an encoded way (Oyama 2001).
The succession of developmental stages from less to
higher differentiated forms rekindled the specter of Lamarckian recapitulation because
of the fear that phylogeny may be understood in progressive teleological terms.
Nonetheless the understanding of ontogeny as a non programmed open ended
process places this fear on shaky ground. There are documented proofs of
The Evo/Devo debate is between preformism and epigenesis that today takes the form of a conflict between the roles to be assigned to genetic information and self-organization respectively. To solve this issue would clarify some lasting discussions that were cast in the following dichotomies: (a) Either the inner nature of the organism or the external environmental conditions as a major cause of evolutionary variations; (b) Individual random variations vs. population statistical regularities.
Waddington's image of canalization of development, as a ball running down through the valleys of an epigenetic landscape whose features are both shaped by a genetic network and the environment, remains a fertile concept for understanding development in its relation to evolution within a more integrative perspective. Genes and environment do not directly cause traits; they rather assist in laying out the main features of the epigenetic landscape and therefore in the pattern of canalizations open to the ontogenetic process. The concept of "genetic assimilation" (Waddington 1957) addresses the question about how specific adaptive genes became fixed, or how was genetic encoding achieved? The epigenetic landscape is a dynamic interface between the genetic endowment of the egg and environmental interactions subject to permanent tuning by organisms that interpret all the informative resources in order to push themselves into developmental pathways while updating their information content (Waddington 1961).
In early stages of development the morphological determinants are epigenetic processes influenced by external forces that act upon primitive cellular aggregates (Ho and Sauderns 1979, Goodwin 1994, Jablonka and Lamb 1995, Jablonka and Lamb 1998, Newman and Muller 2000). These external determinants work as causal factors of development that may be reinforced by chemical compounds genetically specified. So to say, genes are suppliers of basic structural proteins and enzymes but do not exert a direct influence on the resultant architecture. The process of genetic assimilation (Waddington 1961) hooked morphogenesis to a genetic circuit that facilitates the generation of digitally encoded templates for raw materials. Thence genes came in to consolidate previous epigenetic processes (Newman and Muller 2000, Salazar-La Ciudad et al. 2003).
Molecular processes like protein folding (Balbin and
Andrade 2004), RNA folding (
DST search for the laws of phenotype construction, within the context of epigenetic and environmental interactions. Phenotype is the developing organisms’ analog record that cannot be decomposed into separate G and E contributions. P is the structure of an organism that determines moment by moment its way of interactions in an environment in the course of its ontogeny. The phenotype is being shaped in the encounter with its surroundings, so that every organism is realized as a totality in its domain of interactions and relations in its P, and lives in a specific P according to its interactions and relations, (Maturana and Varela 1992, Maturana and Mpodozis 1995). Thus, P changes in the course of ontogeny result of both its internal dynamics and its interactions with E.
The semiotic integration:
I will go in depth into Taborsky’s model (Taborsky
2004) in which both domains ontic (independent of observer’s knowledge) and
epistemic (dependent on observer’s knowledge) are considered simultaneously and
their mutually entailing relations made explicit. Moreover, the acceptance of
living systems as inner observers bridges these two cuts and paves the way for
an integrated view of evolution and
development that highlights the existing parallelisms between them that I list
as: 1. Self-referentiality. 2. Autonomy. 3. Open-endedness (divergence). 4.
Increase of information. 5. No existence of a prior program. 6. Irreversibility.
Most of the issues that concern the Evo/Devo debate boil down to irreconcilable background assumptions grounded on the dominant Cartesian, dualistic, mechanical and reductionist perspectives. A semiotic paradigm discovers a raison d'être beyond the internal/external, individual/population or local/global spatial domains, and present/perfect/progressive time. The six coded modes inspired by Peirce’s categories must be developed and put into the biological discussion as an alternative reference frame (Taborsky, 2002, 2004).
To reiterate – the six relations defined by Taborsky are:
Firstness-as-Firstness [1-1]: Internal local/Present time/Possible (Internal analog).
Secondness-as-Secondness [2-2]: External local/Perfect time/Discrete actual (External analog).
Secondness-as-Firstness [2-1]: Borderline interface/Perfect-present time/Attractor phase.
Thirdness-as-Firstness [3-1]: External global/Progressive-present time/Statistical average.
Thirdness-as-Secondness [3-2]: Internal global/Progressive-perfect time/Future propensity (Digital).
Thirdness-as-Thirdness [3-3]: Aspatial/Atemporal/Imaginary.
These six relations function as semiosic
codal measurements; when selection is made from these six and linked within a
triadic functional set, the result is a morphological reality, a sign. Whether
this morphological unit is open or closed, is fuzzy or crip, is an atom, a
cell, a word, an action – it is a sign, a morphological reality, a
well-formed-formula.
(Taborsky 2004)
This paper sketches a morphogenetic analysis beyond the dichotomy that conceives determinism on either genetic (internal) or environmental (external) factors, and randomness as originating either from within (mutation) or from without (environmental fluctuations). Although, internal/external and local/global boundaries are so thoroughly implicated as to be almost indistinguishable, the analytic dissection of these spatial-temporal-functional zones would permit to examine the relationships between ontogeny and phylogeny.
The reconstruction of the bridges between ontogeny and phylogeny will contribute to an expanded evolutionary synthesis that would be Darwinian in the sense that agrees with his initial interest of explaining divergent evolution in terms of embryonic development.
Ontogeny obeys instructive information, and evolution obeys selective information; the former is commonly associated with directed variation and the latter with random variations (Jablonka and Lamb 1995, 1998). Instructive information is equivalent to analog information (Hoffmeyer and Emmeche 1991, Hoffmeyer 1996) that can be divided into external and internal zones encoded in the [2-2] and [1-1] modes respectively, (figure 1, quadrants I and II). Analog information is defined as direct pattern recognition by some structural motifs of the evolving agent within a continuous threshold of variability. Analog refers to the fact that this recognition is made either by structural complementarity or by similarity and thus, it permits the establishment of non-random reversible interactions (Root-Bernstein and Dillon 1997). If the motif recognized is external, then interactions and measurements lead to internalization of information. If the recognized motif is internal, we have an interaction among inner constitutive components that preserves the coherence of the individual as it interacts with the environment. Digital information refers to encoded information in the form of a record that has the structure of a text composed of basic symbols (DNA) that can be modified by discontinuous variations such as mutation and recombination. Digital information is encoded in the [3-2] mode, which is associated with information that acts as a network that defines future propensities. Genes do not exactly belong to the relation [2-2] because they do not have a defined material boundary; they overlap, they are physically discontinuous as they are intervened by other sequences, they can be found within other coding sequences, their continuity is observed at the RNA level. Genes might be thought of as a functional networked unit under the command of the whole organized epigenetic system that interacts with the environment. Genes as informational sources belong to the [3-2] mode. The discreteness of the genes is a Mendelian approximation but it does not hold at the DNA level. To map Mendelian genetics into molecular genetics is almost impossible.
Digital information appears when any structure
becomes a symbol that can be permuted giving rise to potentially different
functional content. The internal and
external analog zones are connected by the epigenetic inheritance systems (EIS)
(Smith and Szathmary 2001,
Jablonka et al. 1992, Jablonka and Lamb 1998), since an adjustment to an
external factor implies a readjustment of inner states. On the other hand,
selective information, or Darwinian natural selection operates from without and
configures the digital genetic informative record shared by the population. The
survivors are the carriers of a more faithful (though outdated) inner
representation of their environment, that is the survival of the best encoders
of environmental information.
These two dualities Lamarckian (internal/external) and neodarwinian (individual/population) are currently the source of endless “either/or” debates. I will show that semiosis favors a coherent integration that is congruent with more recent developments in Developmental Systems Theory (DST). There is a fundamental symmetry between genes and other developmental resources (Oyama 1985, Griffiths and Gray 1994). The semiotic approach is based on the idea that there is a real interpretation of all informative resources by the developing organisms.
2. Igus As Evolving Developing Agents
The notion of Evolving and Developing Agents (EDA) aims to
formalize a theory of organisms as real interpretants that choose between
alternative ways to relate with external factors and so encode and incorporate
information from the environment. The term merges development and evolution
inasmuch as both processes are mediated by the agency of the organisms
themselves. This notion agrees with Baldwin’s idea of organisms as participants
in their own adaptations by means of organic selection (
The upper quadrants in figure 1 correspond to asymmetric relations while the lower ones account for symmetry or regularities that in Peirce’s view are properties of Thirdness. If we consider the potentiality inherent to digitally encoded information (quadrant III) one can talk of ontogeny as the transformation from the genetic to the phenotypic at individual scale, or from the potentialities given in the present to an already manifested determination (perfect time) that is from [1-1] to [2-2]. However, this transformation requires an interpretant that Hoffmeyer (1996) locates in the egg, since it reads DNA instructions and interprets them to implement an ontogenetic trajectory.
Embryogenesis, or ontogenesis, is a sign operation in the sense that a one-dimensional “DNA inscription” containing a coded version of its parents is converted in a three-dimensional organism… An inscription is not meant to be active, it is meant to be deciphered, and so it is with the DNA inscription. …. The egg – or the growing embryo – is the some one for whom the genome represents a sign of a specific process of development. (Hoffmeyer 1996)
Likewise evolution as a population phenomenon is usually understood as a transformation from the phenotype to the genetic at the level of population by means of an interpretant in this case the lineage.
Any given lineage interprets its ecological niche as a sign that the DNA pool must be modified. (Hoffmeyer 1996)
The interlocking of these two processes is clearly seen in the fact that every single phenotype that makes part of the evolving population proceeds from an individual developmental process, and that individual DNA sequences that are interpreted along development come from a previously selected population (genetic pool). However the complexity of the processes involved requires that the internal/external and individual/population interface encoded in the [2-1] mode be specified. I name this interface as Evolving Developing Agent EDA or an Information Gathering and Using System (IGUS) that acts firstly as a random explorer of the possibilities of the environment through measurement and secondly tends to interact or to promote couplings that configure a stable inner record, because structural adjustments to environmental factors involve an inner structural modification. The ‘thingness’ of the Agent or the System would be evident only within a triadic morphology that involves a relation in the mode of Thirdness (Taborsky 2004). IGUS pick up input information in order to enhance evolutionary adaptability and capacity to promote individual diversity, so opening up future possibilities. The exploration of the environment by the EDA is close to random only to start with, but as information is gathered it becomes more directed or canalized. As I will show below, there would never be enough information to make a directed successful response, EDA are, thus, compelled to make choices or to risk something.
An IGUS is defined as a physical entity that is interested in free energy or extractable work. As an inner observer it interacts, its cognitive activity is error prone, finite, partial and uncompleted, but good enough to preserve its performance and identity. An IGUS was conceived as a universal Turing machine that can extract work taking into account the balance of entropy production as a result of data processing. In order to estimate the amount of free energy that can be converted into useful work, Zurek (1989, 1990) defined "physical entropy" (S). An IGUS can also be imagined as an agent that probes the surroundings by performing a series of measurements and processes the results in order to optimize the amount of useful work, (Kauffman 2000), (Andrade 2003, 2004). As the number of measurements proceeds, its uncertainty about the environment decreases and the gained information increases the size of the digital record (see figure 2).
(Figure 2)
Physical entropy (S) is equal to the sum of two independent quantities: Missing information (H)[1] plus Chaitin´s algorithmic complexity (K) of the string that records the known data. Therefore, for any state of the system S = H + K. An IGUS acts as the converting agent of missing information into known randomness (K) of the record[2]. Measurement decreases the ignorance about surroundings, but increments the size (algorithmic randomness) of the record that encodes the obtained information. The model explains how the amount of energy that can be used by the IGUS is dependent on the algorithmic complexity of the record, since the more compressed the record (the lower K), the higher the amount of work (W) that can be extracted. The difference between DH and DK determines the net gain of work. Far-from-equilibrium, DH is greater than DK, and a quantity of net work can be extracted, by contrast in equilibrium DH would be equal to DK and no net work can be extracted. The net work (DW) obtained at temperature T is given by the sum: DW = DW+ + DW-. Where DW+ is the work gained due to the change of H, given by DW+ = T (Hf - Hi). DW- is equal to the cost of observer's memory updating that replaces the outdated record ri about the initial state, with the record rf that describes his knowledge about the final state, given by DW- = T (Kf - Ki) = T ([r*f] - [r*i]). DW- = Ki/f = r*i/f = [r*i] - [r*f]. Let r*i/f be the size of the minimal program that computes ri given r*f. The extraction of energy becomes possible only through compression of the record (r) that is dependent on efficient encoding, or the IGUS' ability to comprehend his experienced world in term of regularities. But the maximal efficiency in energy extraction can only be attained by the IGUS when ri/f is the minimal program r*i/f, that can only be accomplished when the number of steps used to find r*, tends to µ in conformity with the halting theorem of computation. In order to be able to extrapolate this model to developmental biology, I assume that: (1) H is the number of possible configurations that an EDA can attain by virtue of its structural organization as such. (2) K is the (specificity and stability) uniqueness of the phenotype. (3) Ki/f stands for (Algorithmic randomness of r*i/f) the average number of intermediary developmental states adopted by the EDA before it acquires the mature structure. (3.1) Kf is the structural uniqueness of P. It would be a function of the number of branching decisions that were made along evolution in order to obtain a particular body plan from a more general structure to a more specific one. (3.2) Ki is the structural uniqueness of P at the onset of development or the primitive conformations. Uniqueness refers to the probability distribution of alternative conformations present in a given state. There are two extreme possibilities: each P is equally likely (one G to many P) or a single P is specified with full certainty (one G to one P).
An EDA measures, classifies and registers environmental
features (Andrade 2004, Balbin and Andrade 2004). Measurements reduce EDA's
uncertainty about E (
In reference to the IGUS, the differentiation
between ontic and epistemic levels of description is problematic (Atmanspacher
2002). H (statistical entropy) can be interpreted as an ontic description of
intrinsic structural probabilities that can be attained by the IGUS. However,
at the same time H can be regarded as epistemic for it refers to IGUS’ environmental
uncertainty, the very fact that H diminishes with measurements means that it is
epistemic, (Burbano 2005). In the same way K (algorithmic complexity) is
necessarily epistemic because it refers to what is known by the IGUS, however K
becomes ontic as long as it can be identified objectively as an analog and/or
digital record, P and G respectively. To summarize in a universe composed of
semiotic agents such as living entities ontological chance is nothing but the
epistemological uncertainty of actually existing agents. If Peirce’s thychism or chance is an objective reality that is operative in the universe
(ontic), then the objective existence of semiotics agents becomes a necessary
condition for the understanding of the most fundamental interactions in the
natural world. Chance as a property of Firstness
refuels the evolutionary potential and creative impulse of nature, however its manifestation
and renewal takes place within the continuous, atemporal and aspatial dynamics
of Thirdness.
This problem has reinforced my conviction about the correctness to extrapolate the IGUS concept to organisms for they exist at the boundaries: (a) “Outside/Inside” since it converts external uncertainty into inner knowledge by measurement; (b) “Local/Global” since individual’s explorations of the environment are converted into digital codes shared by the population (c) “Present tense” potentialities/“Perfect tense” actualities because organisms’ ever going actions in the “progressive” time mediate between them.
IGUS refers to objective entities that partially know their immediate environment. They dwell in the ontic and epistemic realms simultaneously as long as they are the organized material structure that results from mind operations and consequently the instrument for further mind operations. Its mindfulness is always in connection to [3-1], [3-2] and [3-3]. Moreover, the internalist perspective, modeled in the manner of a semiotic agent brings together the ontic and the epistemic realms. This heuristics helps to understand the inseparability of subjective and objective reality, because it asserts the existence of organizing agents that act as participants (subjective reality) in a permanent coevolutionary unfolding with their environment (objective reality), since agents shape their environment while are being imprinted by it. In other words, organisms encode information from an environment that is being modified by their activities that exert an impact on it.
The mode [2-1] shows that the interface is always being trespassed in either direction. An IGUS passes from uncertainty (H) to certainty (K) by measurement, while uncertainty (H) increases as a result of either record erasure (decrease in K) or environmental change. Agency refers to the fact organisms regulate and buffer genetic (G) and environmental (E) influences. Structural adjustments in which some configurations are preferred in accordance to specific environmental conditions is one of the distinctive features of cognition. Thus the autonomy of the EDA depends on contingent evaluations and interpretations of both G and E resources that lead to unpredictable phenotypic outcomes. The evaluations and interpretations made by the EDA are beyond what an external objective observer can predict and so constitute the real source of logical non-predictability.
Digital information in the [3-2] mode provides
multiple options through the participation of the EDA that manifests as a
material entity partially closed, partially open, individual in a population
context only when it operates in connection to Thirdness. Organisms are transient manifestations of processes and
relations. All relations collapse into the [2-1] only as a material
organization that tend to individualization, but simultaneously the same relations
remain diffused in the [3-3] mode to the extent that without them individualization
is not possible. Reality is not pure matter, nor pure mind but something that
manifests as an interface in a dynamic relation that as it tends to closure we
identify as [2-1], as it remains open is identify as [3-3], that is why [2-1]
and [3-3] do not fit properly into the Cartesian quadrant. The Cartesian
quadrant is useful for it helps to show this limitation.
The interface [2-1] cannot operate alone. As long as it gets localized it
makes part of the physical material world, but as long as it is part of the
population it interacts with the relations [3-2] to facilitate work extraction.
The captured free energy connects again with the [1-1] relation. The relation [2-1]
is able to establish interactions to construct the network, while interactions
based on [2-2] are only mechanical and do not contribute to the network of
information exchange and processing. IGUS accounts for network expansion
because they are partially open, the isolated closed entities in the [2-2] mode
cannot do that.
Thus, an EDA must be studied simultaneously in two time-frames, ontogenetic and evolutionary. In both cases, the record reflects its knowledge about the state of the system. [Note: From here onwards, I will refer to the IGUS exclusively as EDA.] The ontogenetic (analog) record is given by the phenotype and it has two measurable parameters: (1) Specificity (how specific is its relation with a particular environmental factor); (2) Stability (how constrained is the span of possible conformations). These two parameters point to the uniqueness of every individual phenotype.
However, EDA’s activity in ontogeny defines and canalizes tendencies that can be verified in evolutionary time. Global long range evolutionary tendencies are the result of local short range processes that occur at the individual level within the ontogenetic context. The key concept of recapitulation theory is that free living young adults of ancestral life forms determined tendencies of change influenced by the environment through change of behavior and habit, and some of these modifications as they were pushed back to earlier ontogenetic steps became part of a descendant species Bauplan. The nature of the evolutionary record is dual: (a) Analog – that refers to evolutionary information that can be inferred from the sequence of ontogenetic paths; (b) Digital – that refers to evolutionary information that can be inferred from DNA sequences.
The duality replicator/interactor (Hull 1978, 1980) evokes the difficulty to split what is ontic from what is epistemic, replicators are ontic since they are entities that pass on their structure by copying a template, and epistemic since replication is done by means of interaction with their environment. In the case of the EDA, the intimate association between these two aspects is one of the consequences of their information processing activity. Like the genotype/phenotype duality, the evolution/development duality is a specific case of a more general analog-digital code duality (Hoffmeyer and Emmeche 1991). In the same vein, to be a replicator implies to have generated a digital record that can be copied, but the generation of a digital record is an analog operation. Beyond this duality we find a deeper underlying unity: Analog-Digital-Semiotic work-actions (Andrade 2002, 2003).
3. Self-Referenciality Is An Intrinsic Property Of Genotype/ Phenotype/ Environment Interrelations
… all life is founded on self-reference. … Would the organism not then be regarded as the DNA analog’s self-description rather than the DNA’s being the organism’s digital self-description?. (Hoffmeyer 1996. Pp. 42 and 44)
Continuity, as generality, is inherent potentiality, which is essentially general. (…) The original potentiality is essentially continuous or general. (Peirce C.P. 6.204-205)
The Weissman’s barrier and the central dogma prevent the influence on the G by the P during individuals’ life cycle, but not necessarily at the population evolutionary time where selection on P determine the genetic pool (G) for the next generation. In addition the fear of violating the central dogma should not stop us from acknowledging that during individuals’ life cycle there may be gain of information from the environment expressed in modification of analog Epigenetic Inheritance Systems (EIS).
A theory of organisms as EDAs or information
encoders and niche constructors, falls into an unavoidable self-reference
circuit because the observer (the interacting P) is included into the digital
(G) description. Organisms participate in the creation of the environment that
acts as an informative source for them, so to say that P as a natural observer
participates actively in the production of both G and E. Both together G and E
partially specify the ontogeny of their offspring, in other words, as
development goes on, the P in ever going consult with the E participates by
tuning and coordinating gene expression throughout the ontogenetic trajectory. Organisms through niche construction and interactions
among them actively participate in the formation of their own inner experienced
environmental representations or Umwelt, so accounting for an
unavoidable non-predictable self-referential loop in the relation organism/environment.
According to Rosen (1990) non-predictable statements are circular
because what is defined is included in the definition. In other words,
something is not predictable if it can only be defined by making reference to
the context or totality it belongs, a totality that can only be defined by
specifying the object we want to define to start with. Circular thinking contributes
to intelligibility but it certainly does not favor predictability. Living
entities are a particular case of non-predictable systems whose behavior is partially
reflected by P/E, G/P and G/E dualities.
Circular thought was eliminated by asserting that development depends on a preexisting genetic program or by affirming that the information flux goes only in one way departing from DNA; however the attempt to solve the questions about the origin of genetic information highlights the need to consider the influences from environment on the phenotype and from the phenotype on the genotype. Similar to the origin of body patterns, the origin of adaptations, the origin of evolutionary variations and in general the major evolutionary transitions are undeniably influenced by the environment in two senses, as a direct contributor to the form by means of physical forces, and indirectly through organisms’ needs to cope with environmental uncertainty.
The neodarwinian approach to evolution and the central dogma of molecular biology did away with closed causal loops in favor of a linear reasoning. Development and evolution make part of a mutually entailing loop that refers to the transformation of life forms. An EDA is explained in terms of its information processing agency that defines the interactions within the same level of organization and the relations with the adjacent lower and higher levels (Andrade 2003). EDA self-organizes by profiting the potential derived from the interpretation of genetic and environmental informative sources. Evolution and development are non-linear processes that take place in a moving trajectory that unfolds within a continuum of different spatial and temporal dimensions, but in order to discern whether a process takes place within inner/external, individual/population zones, it is desirable to identify for each transition the mode in which analog and digital informational processes overlap. The continuity of these natural processes is given by the [3-3] mode, a mode that was anticipated by Aristotle when he referred to the continuity of causal agencies. Considering nature as a developing process that actualizes form, the formal cause or form is the cause of movement because it mediates between final and efficient causes, (Aristotle. Phys. III, 1). The efficient cause (source of change) is a form operating a tergo, and final cause a form operating a fronte. In many cases form, source of change, and end, coincide because when a form is a source of change, it is a source of change as an end. In other words, Aristotle implicitly states the need to close the circuit from Final to Efficient causality, but this circuit is necessarily beyond formalization.
4. Edas Execute Measurements In Order To Cope
WITH
ENVIRONMENTAL UNCERTAINTY
My aim is to substantiate the following propositions:
1)
Ontogeny and evolution are
aspects of the same General
Transformative Tendency.
2)
This General Transformative
Tendency is driven by EDA interactions with surrounding elements that result in
a functional compromise between: (a) Increments of the phenotype’s uniqueness
(stability and specificity), and (b) Anticipation of environmental changes.
3)
The elimination of redundancy
or the minimization of developmental steps must be positively selected.
This general transformative tendency describes a vector of an inherent driving force to increase mutual information content between Phenotype/Environment that can be decomposed into two vectors: developmental and evolutionary. The former describes increases in Genotype/Phenotype mutual information content, while the latter describes increases in Genetic/Environmental mutual information content.
(Figure 3)
An EDA interacts in order to acquire and to process
information that is used to modify the strategies of free energy extraction
from surroundings. Measurement is any kind of interaction between an observer
system and an observed one that requires classification of the latter by the former
(Pattee 1995). The process of classification narrows the initial set of
alternatives or choice of possibilities. Measurement is logically irreversible
because it establishes a mapping of the kind "many to one" (Pattee
1967), and so generates a simplified and functional observed system's
description (Pattee 1995). In other words, classification is a process in which
the relevant elements of an observed system are selected by the observer that
neglects those that are unimportant. An event of measurement requires two
steps: (1) Recognition, (2) Adjustment. Recognition is mediated by structural
complementarity that uses a structure as a measuring standard of classification
used to compare a variety of external factors. Recognition induces structural
readjustments that narrow the set of conformations, so that the number of different
individual conformers for the uncoupled state is higher than for coupled
states. Interactions reduce phenotypic plasticity inasmuch as the specificity
and stability of P depends on environmental conditions. Phenotypic adjustments
register a simplified functional environmental description that corresponds to
the set of configurations for the coupled state; this is, thus, the record of E
as it was recognized by EDA. Recognition is evident in the interaction between
EDA and its E in both evolutionary time and ontogenetic time. Therefore, EDA decide
based on measurement outcomes since their autonomy and activity depend on the
gathered information from E. Encoding efficiency is a
function of the ability to "model" the experienced world in terms of
regularities.
Phenotypes are the actual interacting structures that lack evolutionary memory and are sensitive to external influences that can affect their own stability. Since they are open to perturbations by E, G encoded information becomes a decisive stabilizing factor of P. EDA permanently strive to cope with E by means of reciprocal adjustments and so creates an interacting network, in which mutual information content between P and E tends to increase while retaining a wide span of fluctuations and flexibility expressed as conditional information. Like individuals living in like conditions tend to act in a similar way, and this tendency defines the range of deviations from the habitual norm produced by the uniqueness of P interactions. Though the adjustment phase is still reversible (in the sense that the character can still disappear when the E stimulus is removed), it becomes a selective condition for genomic variants that fix the record in an irreversible encoding. Organisms produce unforeseen E perturbations as they try to cope with E, thus, the increasing inner coordination makes it more capable of dealing with a wide range of E changes.
The fact that developmental processes can be both extended by addition of terminal stages and condensed in the sense that features tend to appear earlier than they first came up explains why the recapitulation of basic patterns of organization is observed as one focuses on processes of growing complexity (Arthur 1997, 2002, Ekstig 1994). However it cannot be expected to take it literally as if every step in phylogeny should appear in ontogeny as a linear succession of stages since besides terminal additions, there may be also terminal deletions, intermediate additions (insertions), intermediate deletions, repetitions, inversion, regressions, etc. In all instances changes that affect late stages of development are more likely to be selected for they do not affect the entrenched early stages of development. The point is that developmental pathways are subject to two seemingly opposite forces that result from EDA semiotic activity: (1) Thirdness manifests as a compressing or shortening force that removes redundant developmental steps without affecting functionality. Also Thirdness is responsible for condensing the time required for each developmental step. (2) Firstness is the expansive force that tends to add new developmental steps and randomize existing structural modules. The joint actions of these two forces manifest in EDA’s semiotic choices or unpredictable adjustments that generate context dependent developmental modifications that account for departures from strict recapitulationism.
The results of EDA’s activity are tracked along the interface individual/population since the P is the record (K) generated at a twofold time scale, in ontogeny, it characterizes the individuals; and in phylogeny the species (the type). The individual record is represented by its P; whereas the population record is given by the probability distribution of the attainable P for every individual that depends on the specific adaptive landscape. Moreover, individual interactions (measurements) change the population record by incorporating environmental information, which modifies the probability of EDA’s accessible states. EDA modifies the adaptive landscape as it evolves, and at its turn the adaptive landscape constraints the possible available P accessible to the individual by evaluating the population’s performance in the environment. On the other hand the interaction with some environmental factors can modify the individual EL and defines the new states available for the population. It is in this sense also that EDA are located at the inter-phase individual/population. The EL is a product of the individuals’ activities where new developmental and evolutionary pathways are opened up.
Development and evolution are the non-programmed process of successive configurations from general to specific that streamlines the production of individual P. The number of states that can be accessed diminishes under stabilizing environmental conditions. As well, measurements bring forth an increment in uniqueness. It is always possible to smooth the passage from one P to another by means of their interaction with external factors. This reduction in roughness of the EL speeds up the ontogenetic path. Since ontogeny involves analog information, shortening of developmental paths (compression of Ki/f) can only be accomplished in evolution if the presence of the surrounding factors is guaranteed. Internalization of these E factors in the form of a symbolic representation accomplishes a state of structural uniqueness while simultaneously the number of neighboring conformations dynamically connected in a morpho-space may increase.
The reduction of redundancy is mostly achieved by taking over a functional redundant structure in order to execute a new task. By this means, an EDA may augment accessibility to new neighboring tasks and reduce ontogenetic time simultaneously. The argument goes as follows, the increase of the fraction of dynamically connected P improve the memory updating efficiency by the EDA (Ki/f), thus, tendencies to gain stability and to improve energy extraction are simultaneously fulfilled. As a result there are observed increments of structural uniqueness. The P is a stationary state within a defined stability threshold of energy barriers (Andrade 2000), work must be done in order to overcome energy barriers in order to attain more stable states, that is to shorten K and to increase uniqueness. As the organism rolls down the EL, it becomes more restricted, canalized, while new ever shallower routes branch down the road, so producing individual intra-specific variations. Consequently its chreod down the epigenetic landscape will run deeper; a fact that makes ever more difficult further shortening of K. In other words, EL was shaped as some P was retained for functionality, stability and uniqueness, so creating deep chreods that drive ontogenetic pathways on its own. It is as if the EDA itself demarcates the path as a truly semiotic agent. As a result of this activity EL are constantly in the process of shaping, the deeper chreods correspond to shortened descriptions (high structural uniqueness, and fast developing), while the shallow valleys correspond to unstable redundant descriptions (low structural specificity and stability or plastic phenotypes that accept developmental changes). To summarize: EDA’s act as a formative cause that identifies environmental regularities and patterns, some of which may be encoded in a compressed manner in two ways: analog (reduction of intermediary states to achieve P), and digital (genetic non redundant description).
This hypothesis provides a unifying explanation for
the following observations and arguments: (1) Primitive life forms consist of
few developmental stages, and fluctuate among different conformations, so that
additional steps were required in order to acquire a more stable form. (2) The
elimination of redundancy streamlines development and increases structural
uniqueness. (3) There is a strong tendency to diversify configurations in early
stages of evolution by the terminal addition of developmental steps. (4) The
evolution of Hox genes has proceeded from dispersion to clustering while
increasing in number, their co-linearity found in most vertebrata expresses the
tendency to optimize encoding.
5. Development And Evolution Are Processes Of Information Increase
According to the foregoing discussions EDAs produce an analog record that is being permanently updated and leads to increasing values of mutual information content, H(organisms:environment). Analog records are the condition for the creation of digital record that register ensuing changes in mutual information content between genotype and phenotype H(genotype:phenotypes) and between genotype and environment H(genetic:environment).
The phenotype/environment relation:
Following Wagensberg (2000), an EDA tends to free itself from the uncertainty of the rest of the world and in so doing reacts to E fluctuations. The point, however, is that the ability to anticipate environmental fluctuations is related to increasing degrees of internal coordination and plasticity. The organism’s striving to cope with environmental challenges originates a tendency to increase mutual information content between P and E, if this information is incorporated into the genetic record, higher mutual information content between G and P, and G and E would observed. While P adjusts to E at the level of the individual, the relations G/P and G/E are detected in the population. The relation P/E is mediated by the organism’s behavior, modeled as an open cognitive system. This is a permanent ongoing functional interaction (progressive time) that produces structural adjustments or analog encoding since to adjust to an external environmental stimuli implies rearranging an internal state.
In other words, every ontogeny as an
individual history of structural change is a structural drift that occurs with
conservation of organization and adaptation. We say it again: conservation of autopoiesis
and conservation of adaptation are necessary conditions for the existence of
living beings; the ontogenic structural change of a living being in an environment
always occurs as a structural drift congruent with the structural drift of the
environment. This drift will appear to an observer as having been “selected” by
the environment throughout the history of interactions of the living being, as
long as it is alive. (Maturana & Varela 1992)
Let us consider EDA’s phenotype (P) and Environment
(E) as informative sources. According to
(1)
H (P) – (2) H (P|E) =
(3) H (E) – (4) H (E|P) =
(5) H(P:E)
Let H(P) (1) stands for diversity of phenotypic states accessible to the organism independently of E. H(P|E) (2) stands for conditional information or the variability of EDA when E is fixed. It refers to phenotypic plasticity or EDA’s anticipatory capacity to E changes. It is the condition for evolvability. H(E) (3) stands for the diversity of states accessible to E. H(E|P) (4) stands for environment sensitivity, impact, or capacity of being changed by EDA. It corresponds to variability of E when the P is fixed. H(P:E) (5) stands for structural complementarity between P/E or the information that circulates between organism and environment. It is the condition of the existence for living entities.
The higher H(P|E), the higher the independence to E uncertainty; the fit organisms/environment is expected to be somewhat loose in order to maintain evolvability. In a fixed environment (low H(E)), individual independence is obtained without any need to increase H(P|E) and specialization becomes an advantage. By contrast in a changing environment (high H(E)), H(P:E) lowers and the systems may either go extinct or conversely make a semiotic election that opens up a new developmental and evolutionary path. EDA achieves independence by investing energy every instant, thus allowing information exchanges with the surroundings in order to maintain the constraints that delimit the interface internal/external. A state of equilibrium is characterized by the absence of constraints between the inner and outer regions, a situation that occurs when the individual is isolated from matter and energy transference. Therefore, an isolated system (individual or population) cannot be influenced from without and changes until it can no longer change in equilibrium. By contrast, every EDA tends by its own activities to remain far from equilibrium. EDA independence is paid by exporting to the outside the effects of its inner activity, so that the ensuing increases of E uncertainty manifest as selective pressures that forces it to change the constraining values at the interface internal/external. As E uncertainty increases, the viable EDA increase their phenotypic plasticity or capacity to choose and so produce further unpredictable environmental change. In consequence the need to cope with E uncertainty is the developmental and evolutionary potential that promotes growing differentiation.
Evolution and development proceeded from primitive low uniqueness P to present day stable and specific P, by means of the acquisition and internalization of environmental information. The plasticity of primitive life forms was buffered by environmental information that shaped the native structures so reducing H (uncertainty), but as evolution went on plasticity was lost due to genetic encoding of some phenotypic features that were originally produced by organisms’ response to environmental challenges. The discrimination of surrounding’s factors is brought about by coupling or complementary interactions between EDAs’ structural modules and environment, thus, EDA enhance its discriminating ability by means of the diversification of its structural modules, (incrementing H(P) and H(P|E) values simultaneously). In other words, variability enables to recognize more features from the environment, thus making it more predictable for the EDA. Therefore, the increase in diversity is needed in order to obtain a more stable and unique P. Remarkably, the transformative drive of the EDA is to increase structural uniqueness by means of creating conditions for the incorporation of variations that are congruent with these tendencies.
There will always be environmental potential information not relevant to the organisms that could be made part of their Umwelt as a consequence of new explorative behaviors that may become encoded in the form of habit. Besides, an EDA may also give up some of its individual identity in exchange for increasing independency to E uncertainty by increasing its dependence on other similar individuals leading to the emergence of a more complex level of organization. For instance the development of multicellular organisms may have started by cooperation among individual cells as a way to decrease individuals’ E uncertainty (Wagensberg 2000).
To summarize: (1) EDA strives to incorporate environmental information as a way to increase phenotype uniqueness. (2) Development and evolution do not depend solely on adaptation to a stable environment, but on independence from a changing environment by maintaining the organisms’ ability to make a semiotic choice. (3) In ontogeny, information increases are detected as a modification in developmental pathways mostly by the addition of terminal steps. (4) In phylogeny, the increases of information are due to selection for phenotypic or developmental plasticity.
The genotype/phenotype and genotype/environment
relations:
An individual EDA acts within a network of EDA(s) that are part of the E. The point is that the mode [2-1] cannot be dealt with in isolation for it is only the material expression of an acting mind in the [3-3] mode. The interface (P, EDA, IGUS) is active, not a passive boundary.
Increases in mutual information content between P and E are the prior condition for further increments in both mutual information content between G and P, and G and E. Specifically, the tendency to increase the content of mutual information between the digital record (DNA sequence or G) and the analog record (structure or P), H(G:P), is the means to enhance the autonomy of EDA in constant E. As it was proposed above, structural uniqueness rises at the interface individual/population that permits the genetic assimilation[3] of innovations in evolutionary time that convert analog into digital information (Andrade, 2002, 2003, 2004). The digitalization of information manifests as increases in H(G:P) and H(G:E) that depict a growing function of time that lags behind H(P:E) changes. In short, the uniqueness of the P was evolutionary achieved by internalization of information from the E in the form of modifications of developmental routes. The modifications that are more likely to arise, affect terminal steps (like additions, deletions and substitutions) because they are less likely to affect inner coordination, although duplication of intermediary steps, and permutation may also be congruent with E changes. These processes, in as much as they contribute to streamline ontogeny, exert a pressure to minimize the increment of G (digital record) as well. Thence, an EDA becomes more dependent on its history and less dependent on the E to the extent that H(G:P) and H(G:E) was internalized in evolutionary time into the genetic record.
To summarize: The relations G/P and G/E are mediated by genetic assimilation. Genetic assimilation explains the incorporation of environmental information into the genetic record. The evolution of ontogeny is a process of information gain expressed as increases of mutual information content between G and P, though P never gets to be fully determined by G. Likewise phylogeny is a process of information gain expressed as increases of mutual information content between G and E, though E information is only partially incorporated into G. Equation (1) can also be applied to understand these relations: H (G) – H (G|P) = H (P) – H (P|G) = H(G:P) and H (G) –H (G|E) = H (E) –H (E|G) = H(G:E), where H(G) stands for genotypic complexity. H(G:P) stands for genetic information that contributes to the phenotype. H(G|P) stands for genes that are not informative for P construction. Conditional information reflects the flexibility of G with regard to P, for instance the capacity to include neutral mutations and adopt new encoded functions. H(P|G) stands for features of P that are not dependent on G, because they are influenced by E. H(P) stands for phenotypic complexity.
Likewise, H(G:E) stands for environmental information that has been incorporated into the genome. H(G|E) stands for anticipatory capacity based on the existence of duplicated redundant sequences. H(E|G) stands for E features not recorded by EDA, though some of them are likely to be recorded in the future. H(E) stands for environmental complexity. Noteworthy all these relations are mediated by the EDA that lives in its phenotype encoded in the [2-1] mode.
The study of the interrelations between G, P and E explains how the evolution of life cannot depend on sheer random variations, but on the joint action of internal/external and individual/population factors that converge to give a more or less directed response based on a semiotic choice that minimizes risks. To assert that all variations are random is to reject the very possibility of evolution, on the other hand to say that all variations are directed means that evolution is fixed and preprogrammed where no original innovations take place, no room for freedom and creativity. Both mistakes are associated with a mechanical view of nature. Chance is ignorant, variations are the very products of semiotic choices neither directed, nor random but canalized. This is a model that minimizes chance without falling into the opposite mistake, a goal directed evolution. Chance is only the unpredictability for an external observer of EDA semiotic choice. Choices are made within a somewhat restricted span of possibilities but which one will be selected depends on EDA’s evaluations. But this choice brings forth new possibilities that were unforeseen before the choice was made. At this moment potentiality [1-1] is renewed.
6. Peircian Relations And Genetic, Epigenetic, Behavioral And Environmental Informative Resources
An EDA experiences and probes its world as an individual that captures energy by encoding and decoding analog and digital information, so pulling itself through a developmental trajectory. The understanding of living entities as EDA becomes an alternative to genetic reductionism, by means of which Developmental systems theory (DST) can be interpreted. Oyama defines a developmental system as a “mobile set of interacting influences and entities” comprising “all influences on development” at all levels (Oyama 1985), and Griffiths and Gray (1994) affirm that it involves a whole matrix of resources and interactions that permit the reconstruction of ontogenetic and developmental information in each generation. Likewise, I propose that the fundamental unit of evolution is not the gene, but the interacting EDA that by measuring and recording is responsible for information updating in each life cycle.
But, to what
extent does the EDA depend on self-organizing processes driven by analog
interactions? And to what extent does the EDA depend on inherited encoded
digital information? To answer this
question one must recall Salthe’s developmental stages, (Salthe 1993: 9, Salthe,
1995): (A) Immature, (B) Maturity and (C) Senescence. Each stage can be
characterized by the relative weight attributed to each one of the six modes of
codification.
A) Immature stage of “openness”:
This phase is suited for modeling the major evolutionary transitions i.e. the origin of life, the origin of the eukaryotic cell, the Cambrian explosion. However at the very earliest before the emergence of life the distinction organism/environment has no meaning. This is the [1-1] mode of full potentiality. It represents all the possibilities at the point where no interactions have yet taken place since the symmetry and homogeneity of an excited undifferentiated matter has not been broken. Nonetheless it helps to understand the burst of diverse and disparate forms achieved in early life history, with no requirement for differential fitness or competition. Once the boundary inside/outside was achieved, life and the environment came into being. The [1-1] mode codes all possibilities of the individual given its inner drives, creative urges and the available energy. For a protein, it corresponds to the state of maximum uncertainty given by the set of all possible attainable conformations from the unfolded state. For an undivided egg, it would be the set of all possible differentiated cells that can be produced by divisions and differentiation, the starting point of a life cycle from unicellular stage towards multicellular organisms. For a primitive life form, it stands for all the possible states or forms that can stem off from it.
Characteristics of phase A:
1.
It is induced by an environmental catastrophe; in consequence
the emerging EDAs are uncertain about their E, a situation that corresponds to
a minimum of shared informational content between EDA and environment and a
maximum of conditional information.
2.
Low connectedness is found between inner components, and
between emerging EDA and E factors.
3.
Non-random interactions are being established between inner
constitutive elements.
4.
Outline of the very basic Bauplan.
5.
Extrapolating Zurek’s model this phase corresponds to high
uncertainty (H) and low digitally encoded environmental information (K). (Fig.
2 stage A).
6.
For evolved life forms evolutionary
innovations are very unlikely in this phase for it would affect early
developmental steps, but if successful, would lead to abrupt intense changes.
7.
The following modes of encoding
overlap: [1-1], [3-3], [3-1].
The asymmetric expansive and creative impulse [1-1] is offset by the habit taking mode [3-3] that through interactions with environmental resources forces a statistical average [3-1] that opens the way for the creation of an interface [2-1].
Early immature stage: prevalence of internal analog information [1-1] with increasing potential for efficient analog/digital conversion. At this stage environmental information has not yet been incorporated into the genetic record, thus, it is a phase of a strong structural dependence on external circumstances, for instance, forms being shaped by external physical forces. D’Arcy Thompson (1942) argued that the basic living forms can be obtained by the action of physical forces from the environment and not by the action of natural selection or even less by the exclusive action of formative inherited factors (genes). The neodarwinian and Mendelian modes of shaping life had to wait for the appearance of encoded information (Newman and Müller 2000). Organization patterns must be inferred prior to the action of natural selection (Kauffman 1993, Goodwin 1994, Denton et al., 2003). In evolution, this phase corresponds to chemical prebiotic evolution, for instance Fox’s micro spheres and Kauffman’s first autocatalytic networks before the emergence of DNA and genetic code.
Late immature stage: it is defined by the appearance of an internal digital redescription of the self in the form of a genetic record in evolution so that thereafter the EDA interprets genetic messages and makes use of gene products. P has a low dependence on G, and the internal analog information plays a role as a cohesive internal factor. There is an abrupt increase of H(P:E) followed by slowly rise of H(G:P) and H(G:E). Associated increments in H(G) bring forth the augment of structural motifs of the EDA that by conferring enough internal variation permits further internalization of information about the transformed environment.
Embryos, thus, correspond somehow to earlier life
forms that were and are no longer open to the influence of external
environmental factors, in agreement with the view that the modification of
developmental routes in free living primitive forms opened up novel evolutionary
paths. It is in this sense that ontogenetic variations create long term
phylogenetic canalizations.
… metazoan development reflects a legacy
of past interactions between variant cell lineages arising within the ontogeny
of ancestral forms. … The fact that metazoans develop via a complex of
epigenetic interactions between cell lineages is prima facie evidence
that the principal modes of metazoan development arose as variants in the
course of ontogeny … Ontogeny must re-enact the interactions which gave rise to
it. (Buss 1987)
Although
B) Intermediary mature stage between “openness and
closure”.
This phase on a developmental scale corresponds to the realization of the juvenile and young adult form and on an evolutionary scale to the consolidation of basic body patterns (Ediacaran explosion of body plans followed by a second explosion around the base of the Cambrian 545 my) and functional adaptation in populations. Maturity is characterized by active encoding of digital information by means of the combined influences of behavior, EIS and cooptation by genetic assimilation of random genetic variations that result in the maximum efficiency analog/digital conversion. At this stage, selection brings forth encoded adapted motifs and the consolidation of the most important structural domains of present day phenotypes.
This phase is characterized by:
1. Rapid increasing mutual information content between P and E that drag subsequent increases in H(G:P) and H(G:E).
2. Increasing connectedness among inner subsystems.
3. Adaptive evolution.
4. Decreasing E uncertainty to a level in which enough plasticity is maintained.
5. Consolidation and fine tuning of structural Bauplan.
6. Extrapolating Zurek’s model, this is a phase of decreasing uncertainty (H) and low increases in mutual information content between EDA and environment (K). (Fig. 2 stage B).
7. For evolved life forms, evolutionary innovations due to modifications of intermediary stages of development are more likely to occur and lead to less abrupt and intense changes with regard to phase A.
8. The following modes of encoding overlap: [3-3], [3-1], [3-2], [2-1].
The following modes of encoding are found, depending on the ways EDA deal with external constraints [3-1] and the interpretations of the genetic information [3-2], the interface [2-1] shifts towards a state of local discreteness in the form of a truly differentiated entity [2-2]. This phase oscillates between “openness” and “closure”, due to EDA’s measurements that happen as a permanent action in the progressive mode [3-3].
When the EDA moves towards openness it can expect to find fairly adaptive evolution encoded in [2-1] and [3-1] modes characterized by a rapid and continuous coupling between organism and environment. Potentiality remains as long as new behaviors or new manners to interpret the environment are being proposed by EDA, some of which are interiorized in Epigenetic Inheritance Systems. In consequence the following can be observed: 1. Directed behavioral innovations. 2. Refueling of organisms’ drive to cope with uncertainty. 3. Enhanced variability of developmental pathways. 4. Increasing mutual information content between G/P and G/E.
When EDA moves towards closure it can expect to find slow adaptive and nearly neutral evolution given by the prevalence of the [3-2] and [2-2] codes. In consequence one can expect to find: 1. nearly neutral genetic variations and increasing redundancy, 2. diminished capacity to anticipate possible environmental changes, 3. stabilization of developmental pathways, 4. decrease of mutual information content between EDA and environment.
C) Senescence
stage approaching “closure”:
The developmental stage of senescence reveals itself as stabilization and decay. On the evolutionary scale, senescence manifests as an ever slower change of terminal adaptation to a stable environment and increasing neutral genetic variations.
This phase is characterized by:
1. Maximum value of encoded information or mutual information content between G/P and G/E.
2. Decreasing capacity of EDA to cope with E changes, loss of flexibility or capacity to react creatively to external stimulus due to lowering of EDA’s conditional information.
3. Overconnectedness among component subsystems.
4. Neutral evolution and random genetic variations.
5. Extrapolating Zurek’s model, this is a phase of minimum uncertainty (H) and maximum values of K as in (1). (Fig. 2 stage C).
6. For evolved life forms evolutionary innovations by terminal modification of development are more frequent but manifest as minor modifications or gradual transitions.
7. The following modes of encoding overlap: [3-3], [3-2], [2-2].
There is an overlap of the modes of encoding [2-2] that is revealed as increasing closure and discreteness, and the mode [3-2] that manifests as a stabilizing of genetic information. The phase of senescence leads in ontogeny to the following outcomes: i) Individual decay and death after completion of life cycle. ii) Reenactment of the juvenile form: the modifications of developmental routes are possible by opening up to new ways to interpret the external environment by young adults’ creative behaviors. For instance: neoteny (early sexual maturity that makes it unnecessary to continue the whole developmental cycle) may bring forth plastic younger forms that lead to novel ways to explore the environments. These outcomes are paralleled by the following evolutionary outcomes: i) Species extinctions. ii) Modified developmental routes that lead to evolutionary branching. This branching will consolidate through the exploration of variants that become functionally coupled to external influences. Notice that in (i) cases, though the fate is death, individuals while still alive make a contribution to the permanent remodeling of the epigenetic and Umwelt landscape. After death they favor the continuity of life by recycling of components. In (ii) cases, new life forms and lineages may spring forth.
In senescence the dependence on digital information follows increasing closure. The specialization of the realized structures occurs as H(G:P) increases towards maximum. At this phase, G undergoes minor modifications that serve to fine-tuning or terminal adaptation, within an already stabilized genetic network. H (G) shows a tendency towards minimum increments but paradoxically, its growth can only be minimized by incorporating more information from the environment. New measurements result in ever smaller changes congruent with ever diminishing entropy production as EDA approaches equilibrium or the point of minimum extraction of available energy.
7. Conclusions
The G/P/E interactions lie as a substrate that accounts for the relations between ontogeny and phylogeny which are not of cause/effect but rather of mutual entailment. If it were a matter of breaking up this logical loop, as mainstream science has done by placing variation prior to selection, and genetic information prior to form, then using the same logic ontogeny should be considered prior to phylogeny. This priority expresses the fact that ontogeny is the source of individual evolutionary variations and also, it is a process in which a digital text is translated into an analog representation. Therefore one would be forced to assert that ontogeny is the causal agency of evolution. However, considering that the self-referential loop does exist in nature I have preferred to justify the idea of a general transformative tendency that manifest at two different time frames. This model does justice to Oyama’s valuable contribution while making it more amenable to formalization by making the analogy between a Developmental System and an IGUS or EDA. On the other hand Taborsky’s cartography of Peirce six modes of encoding becomes a brilliant philosophical perspective that sheds light on deeply felt scientific problems by providing a way to interpret empirical data. The material embodiment of the EDA is encoded in the analog [2-1] and [2-2] modes but as an agent of interpretation its activity is associated to [3-1] and [3-2] modes of being. Notwithstanding the differences with previous models is that cognition is not a property exclusive to the population/global realm but also to the individual/local, a fact that is best expressed in the idea of the EDA located at the interface internal/external, individual/population, present/perfect.
EDA is the phenotypic attractor state where environmental and genetic information are instantly interpreted, the interaction with the environment becomes recorded in physiological, behavioral and EIS adjustments, that switch on and off existing genes while creating condition for further assimilations of new genes. Newman and Muller (2000) asserted that the relationship between genes and form is a derived condition, a product of evolution, a statement that I hope to have explained in depth by pointing out that it is the outcome of EDA semiotic choices.
This model explains also why there should be a tendency to retrieve developmental programs that are as short as they can be. The presence of non-redundant developmental programs is best accounted by a semiotic process mediated by a system sufficiently complex such as to be able to rid some steps off. This phenomenon becomes also the reason why strict recapitulation is unlikely to happen, though the core of the hypothesis is preserved, that is that novel evolutionary variation arose as organisms’ reaction to environmental modifications in free living ancestors in their juvenile stages through changes in behavior that paved the way for major modifications some of which became genetically fixed. To conclude, natural selection favors EDA that is able to implement semiotic choices.
Acknowledgements
I acknowledge the support of the Department of Biology and the Vicerrectoria de Investigaciones de la Universidad Nacional de Colombia. I am grateful to Edwina Taborsky for thorough reading of the manuscript and helpful discussion and to Hernan Burbano and Carlos Maldonado for comments.
References
Atmanspacher, H 2002. Ontic and Epistemic Descriptions of Chaotic Systems. In:
Between Chance and Choice. Edited by H.
Atmanspacher and R. Bishops. P:7.
Andrade,
E. 2000. From external to internal
measurement a form theory approach to
evolution. BioSystems 57 (2): 49-62.
Andrade, E. 2002. The organization of Nature: Semiotic agents as intermediaries between
digital and analog informational spaces. SEED Journal (Semiosis Evolution Energy
Development) 2 (1): 56-84.
Andrade, L.E. 2003. Los
demonios de Darwin. Semiótica y Termodinámica de la Evolución
Biológica.
Andrade, E. 2004. On Maxwell’s demons and the origin of evolutionary variations: and
internalist perspective. Acta Biotheoretica
52: 17-40.
Arthur, W. 1997. Animal Body Plans, A Study in Evolutionary Developmental Biology.
Arthur, W. 2002. The emerging conceptual framework of evolutionary developmental
biology. Nature 415 (14): 757-764.
Aristotle.1992. Physics (Book I and II). tr. William Charlton.
Balbin, A. and Andrade, E. 2004. Protein folding and evolution are driven by the Maxwell
demon activity of proteins. Acta
Biotheoretica 52: 173-200.
Bowler, P.J. 1992. The Non-Darwinian Revolution. Reinterpreting a Historical Myth. The
Burbano, H. 2005. Determinism, Indeterminism and Semiotic election. (5th Gathering in
Biosemiotics).
Burkhardt, R.W. jr. 1995. The Spirit of System. Lamarck and Evolutionary Biology.
170.
Buss, L.W. 1987. The Evolution of Individuality.
Chaitin, G.J., 1975. Randomness and
mathematical proof. Scientific American
232: 47-52
Darwin, C. 1859. The origin of species. First edition.
Chap. 13.
as the major determinant of biological complexity in the subcellular realm: new support
for the pre-Darwinian conception of evolution by natural law. BioSystems 71: 297-303.
Depew,
D.J. and B.H. Weber. 1995. Darwinism
Evolving. System Dynamics and the
Genealogy of Natural Selection. A
Ekstig,
B. 1994. Condensation of developmental stages in evolution. BioScience
44: 158-
164.
Transitions. Science 280:
1451-1455.
RNA Genotype-Phenotype Mapping. Journal Theoretical Biology 194: 491-515.
explanation. Journal of Philosophy
(16): 277-304.
Philosophy 12 (4): 471-492.
Goodwin, B. 1994. How the leopard changed its spots.
Nicolson.
Haeckel,
E. 1879. Evolution of
Embryos,
Genes, and Evolution. The
Developmental-Genetic Basis of Evolutionary
Change.
Ho Mae Won. and Saunders, P. 1979. Beyond neo-darwinism. An epigenetic approach to
evolution. Journal Theoretical Biology 78: 573-591.
Hoffmeyer,
J and C. Emmeche. 1991. Code-Duality and the Semiotics of Nature. In:
Hoffmeyer, J. 1996. Signs of Meaning in the Universe. Trans. by B.J. Haveland.
Hull D.L. 1978. "A Matter of
Individuality". Philosophy of Science 45: 335-360.
Hull D.L.1980. "Individuality and Selection". Annual Review of Ecology and Systematics
11: 311-312.
Jablonka, E., Lachman, M. and Lamb, M.J. 1992. Evidence mechanisms and models for
the inheritance of acquired characters. Journal of Theoretical Biology 158: 245-268.
Jablonka, E. and M.J. Lamb. 1995.Epigenetic Inheritance and Evolution. The
Lamarckian
dimension.
Jablonka, E. and M.J. Lamb. 1998.Epigenetic
inheritance in evolution. Journal of
Evolutionary Biology 11: 159-183.
Jacob, F. 1982. The
Logic of Life, a History of Heredity.
Pantheon Books.
Pp: 147-148.
Kauffman, S. 1993. The Origins of Order: Self-Organization and Selection in Evolution.
Kauffman, S. 2000. Investigations.
Lamarck, J-B. 1803. Zoological Philosophy.
and reprinted in 1963. Chap VII.
Maturana,
H. and Varela, F. 1992. The Tree of
Knowledge. The Biological Roots of Human
Understanding. Pp:
94-117. Boston, MA. Shambala.
Maturana, H. and Mpodozis Marin,
J. 1995. Origen de las especies por medio de la
deriva natural. In: H. Maturana. La Realidad ¿Objetiva o construida?.
Universidad
Iberoamericana (México). Ed. ANTHROPOS. Pp: 105-159.
Newman, S. A. and Müller, G. B. 2000. Epigenetic Mechanisms of Character Origination.
Journal of Experimental Zoology 288: 304-317.
Oyama, S. 1985/2001. The Ontogeny of Information. 2nd
revised ed.
University Press.
Pattee, H.H. 1967. Quantum
mechanics, heredity and the origin of life. Journal
of
Theoretical Biology 17: 410-420.
Pattee, H.H. 1995. Evolving self-reference: matter, symbols, and semantic closure. In:
Luis Rocha (Ed.), Communication and Cognition - Artificial Intelligence (Special issue
Self-reference in biological and cognitive systems) 12 (1-2): 9-27.
Peirce,
C.S. [1965 ]. 1987. Collected Papers. Cambridge, MA: Belknap Press. Obra Lógico
Semiótica
tr.
and ed. Armando Sercovich. Madrid: Taurus Ediciones.
Peirce, C. S. (1931-1958). Collected Papers of Charles Sanders Peirce. Vols.1-8. C.
Hartshorne, P. Weiss, and A. W. Burks (eds.).
Press. CP 1.409. 1.415. 1.416.
Richards,
R.J. 1992. The Meaning of Evolution. The Morphological Construction and
Ideological Reconstruction of
Raynaud. 1997. Haeckel, embryos and evolution. Science 280: 983-984.
Root-Bernstein,
R.S. and Dillon P.F. 1997. Molecular Complementarity I: the
complementarity theory of the Origin and
evolution of Life. Journal of Theoretical.
Biology 188:
447-479.
Rosen,
R. 2000. Essays on Life Itself.
(Complexity in ecological systems series). Pp: 184.
Salazar-La Ciudad, I., Jernvall, J. and Newman, S. 2003. Mechanisms of pattern
formation in development and evolution. Development 130: 2027-2037.
Salthe, S. 1993. Development and Evolution. Complexity and Change in Biology. Pp: 9,
45 and 181-185.
Salthe, S.N. 1999. Energy,
Development, and Semiosis. in Edwina Taborsky ed. Semiosis,
Evolution, Energy: Towards a Reconceptualization
of the Sign. Pp: 245-261.
Shaker Verlag.
Smith, J.M. & Szathmary, E. 2001.
The Major Transitions in Evolution.
University Press. Pp: 247-250.
Taborsky, E. 2002. The Six Semiosic Predicates. SEED Journal (Semiosis, Evolution,
Energy, Development) Vol. 3 No. 2. pp: 5-23.
Taborsky,
E. 2004. The Nature of the Sign as a WFF – A Well-Formed Formula. SEED
Journal
(Semiosis, Evolution, Energy, Development) Vol.4 No.4. pp: 5-14.
Thompson D’Arcy 1942. On Growth and Form. 2ed.
University Press.
Waddington,
C.H. 1957. The Strategy of the Genes. Geo
Allen & Unwin.
Waddington, C.H. 1961. Genetic
assimilation. Advances Genetics 10: 257-293
Wagensberg, J. 2000. Complexity versus Uncertainty: The question of Staying Alive.
Biology and Philosophy 15: 493-508.
Zurek, W.H. 1989a. Algorithmic
randomness and physical entropy. Physical
Review A.
40 (8): 4731 - 4751.
Zurek, W.H.. 1989b. Thermodynamic cost of the computation, algorithmic complexity
and the information metric. Nature
341: 119-124.
Zurek, W.H., 1990. Algorithmic information content, Church - Turing Thesis, Physical
Entropy, and Maxwell's demon. In: Zurek, W.H (ed.), Complexity, Entropy, and the
Physics of Information, SFI Studies in the Sciences of Complexity, Vol. VIII. pp: 73-89.
Addison-Wesley.
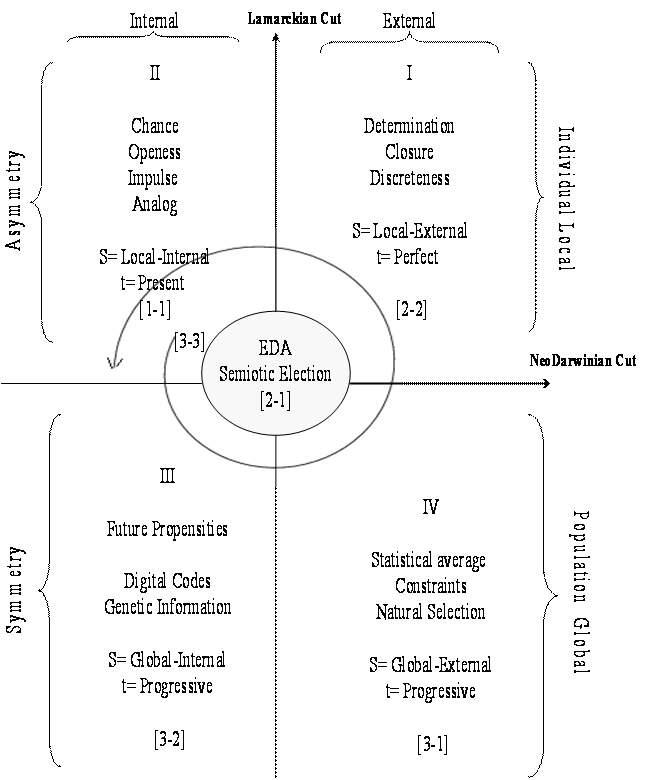
Figure 1. The
Lamarckian/Neodarwinian Quadrant (modified after Taborsky 2004). The Y-axis refers to the
Lamarckian (ontic) cut and the X-axis to the Neodarwinian one (epistemic). The
left upper quadrant (II) corresponds to the internal individual zone [1-1] and
the right upper one (I) to the external individual [2-2]. Development
corresponds to a movement from quadrant II to I by the action of an inner
impulse to differentiate and manifest in a more stable and determined manner.
Evolution as a population phenomenon goes from quadrant (IV) [3-1] to (III)
[3-2], the effect of a stabilizing statistical law that configures an inner
digital record.
![]()
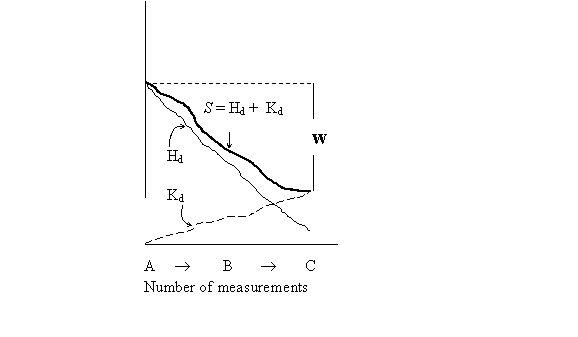

Figure 2. Zurek's
physical entropy S = H + K
variations against the number of measurements performed by an IGUS in a
far-from-equilibrium environment. Bold curve represents the variation of S.
Faint curve represents variations of H with the number of measurements. Dotted
curve represents the variation of K with the number of measurements. W stands
for the amount of work that can be extracted by the IGUS. From left to right A,
B and C stand for one cycle of an evolutionary process. Stage A: Early
evolution / openness / prevalence of H. Stage B: Intermediate stage of evolution / partial closure / maximum conversion
from H to K. Stage C: Stabilizing evolution / closure / dominance of K.
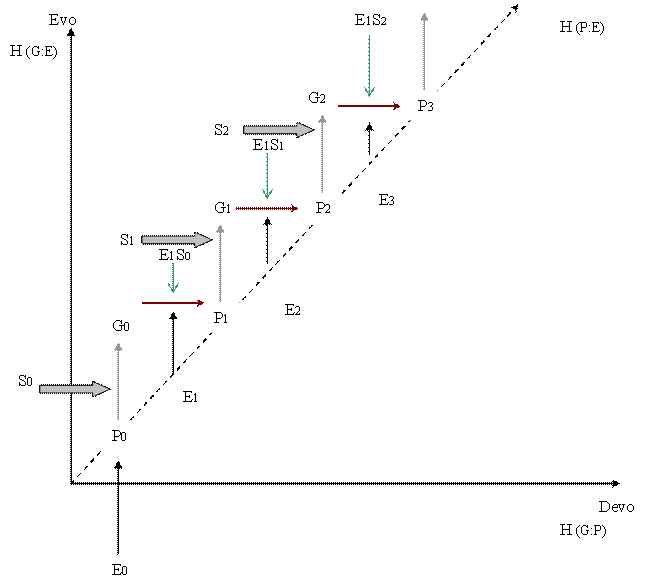
Figure 3. The dashed diagonal shows
increment in mutual information content between P and E, as a permanent action
in progressive time. Pi stand
for phenotypes in which EDA live that go from low to high uniqueness. This
vector can be decomposed into a vertical and a horizontal one. The vertical
axis shows increments of mutual information content between G and E along
evolutionary time. The horizontal axis shows increments of mutual information
content between G and P along the evolution of development. Ei stand
for environmental physical influence on shape that decreases with evolutionary
time: E0 < E1 < E2 < E3. EISi stand for epigenetic
inheritance systems that show a growing tendency along time: EIS0 > EIS1 > EIS2 > EIS3 . Si
corresponds to selection pressure.
Horizontal arrows from G to P stand for development at the level of
individual, while vertical arrows from P to G stand for evolution at the level
of the population.